make pdf && rsync
してから富良野の後藤さんにメイルかき.
他の部分の書きなおしが進捗中のようなので,
当方のトドマツ作業はここでいったん停止する予定.
$HOME
で
make log
するだけで「更新」されてしまって,
ですね.

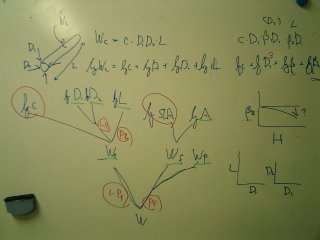
library(lattice) trellis な作図の修行してたんだよね
(「これが我が逃走経路だ!」)
……
とりあえず,
こんなかんぢで
(→ 全体).
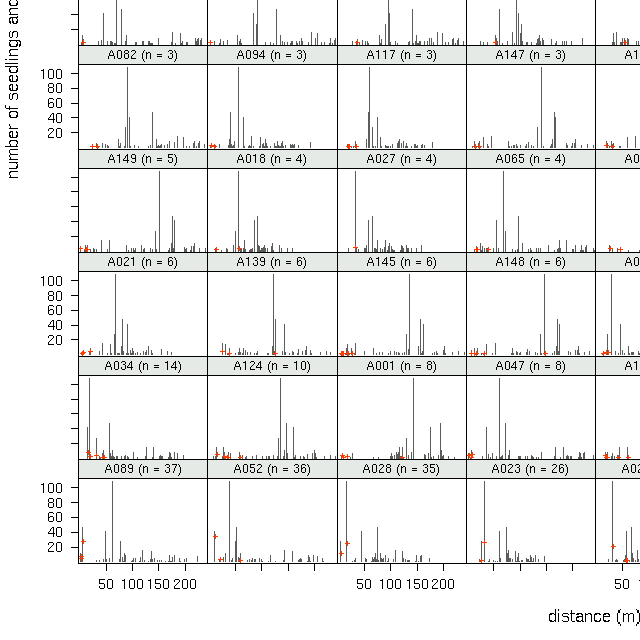
いろいろ書いてて気づいたのですが,このモデルも含めて従来の種子散布や実 生分布モデルのまずいところは「各種子独立」を仮定していることかもしれま せんね.おそらく種子は一個一個が独立ではなく,一個実生がみつかればその 近傍に同じ親からきた実生がみつかる確率が高い,といった空間相関があるの かもしれません.どうもこのトドマツデータはむしろそちらで説明されるよう な気がしてきました.まあ, 「各種子独立ではない」モデル, はまた「次」の機会にしよう.
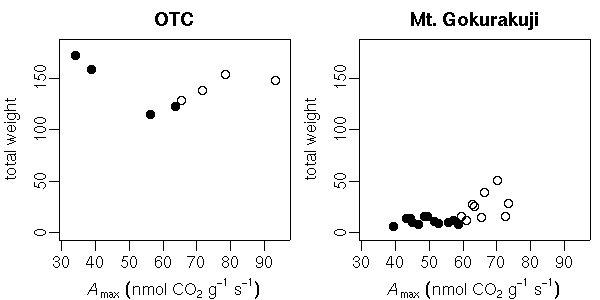
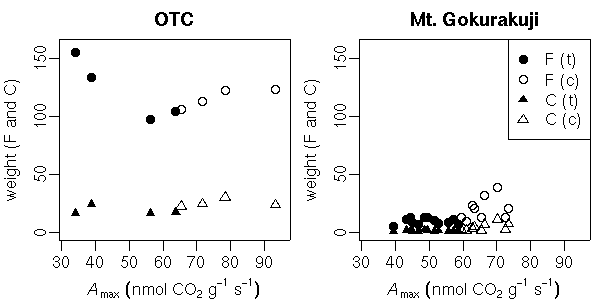
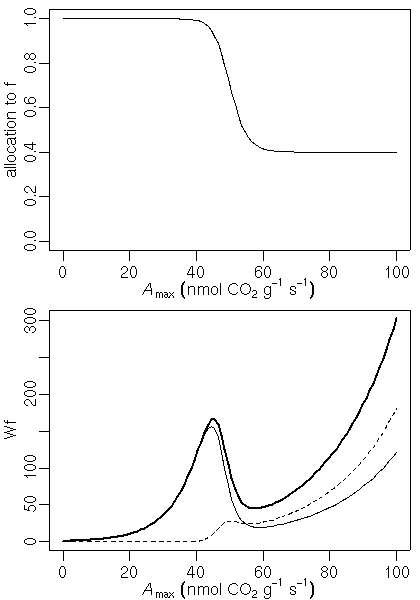
sample() 関数をつかうと簡単にできる.
手順はこう:
-> v.distance
-> v.weight
max(sample(v.distance, ni, prob = v.weight))
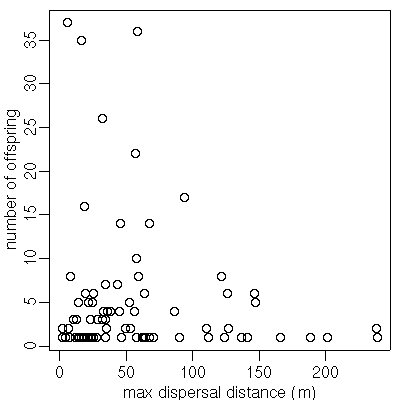
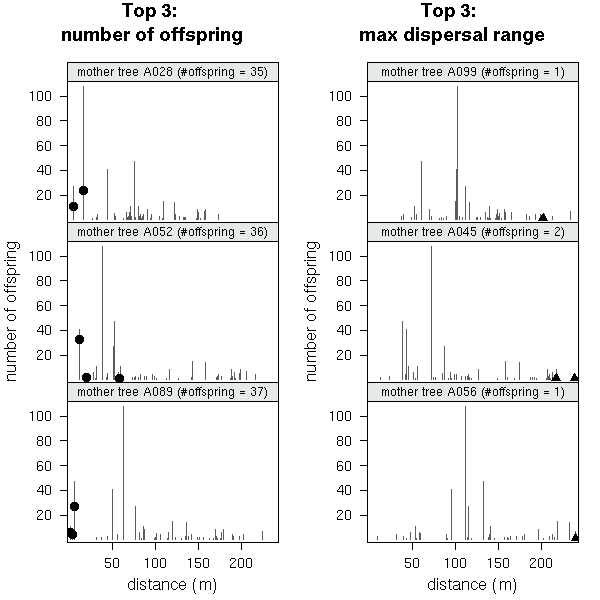
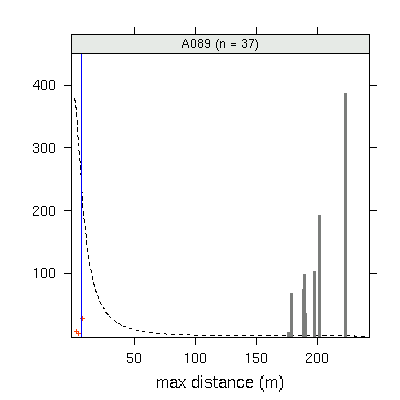
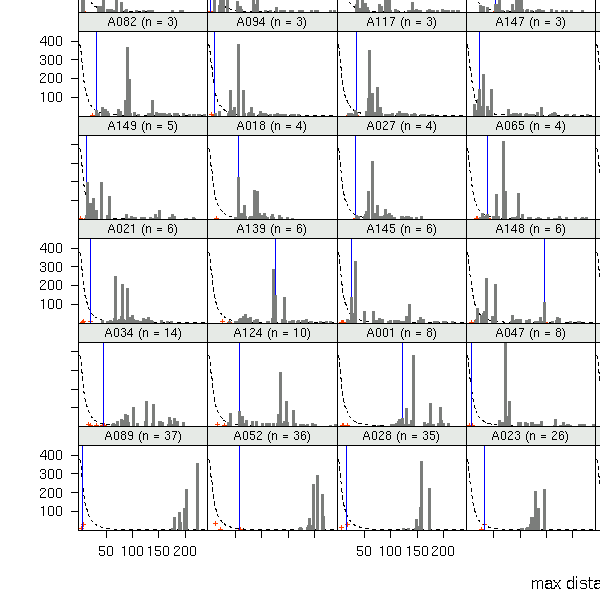
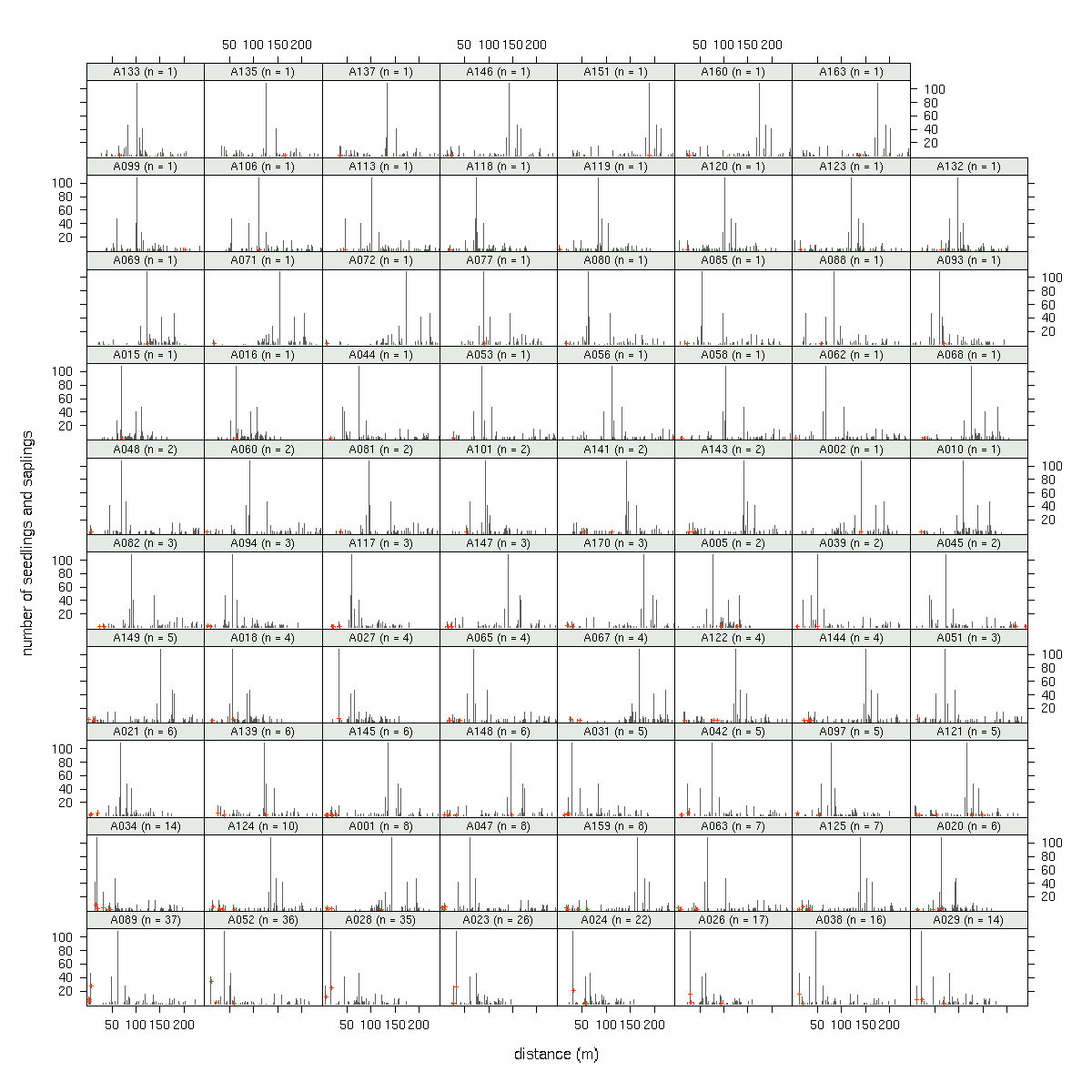
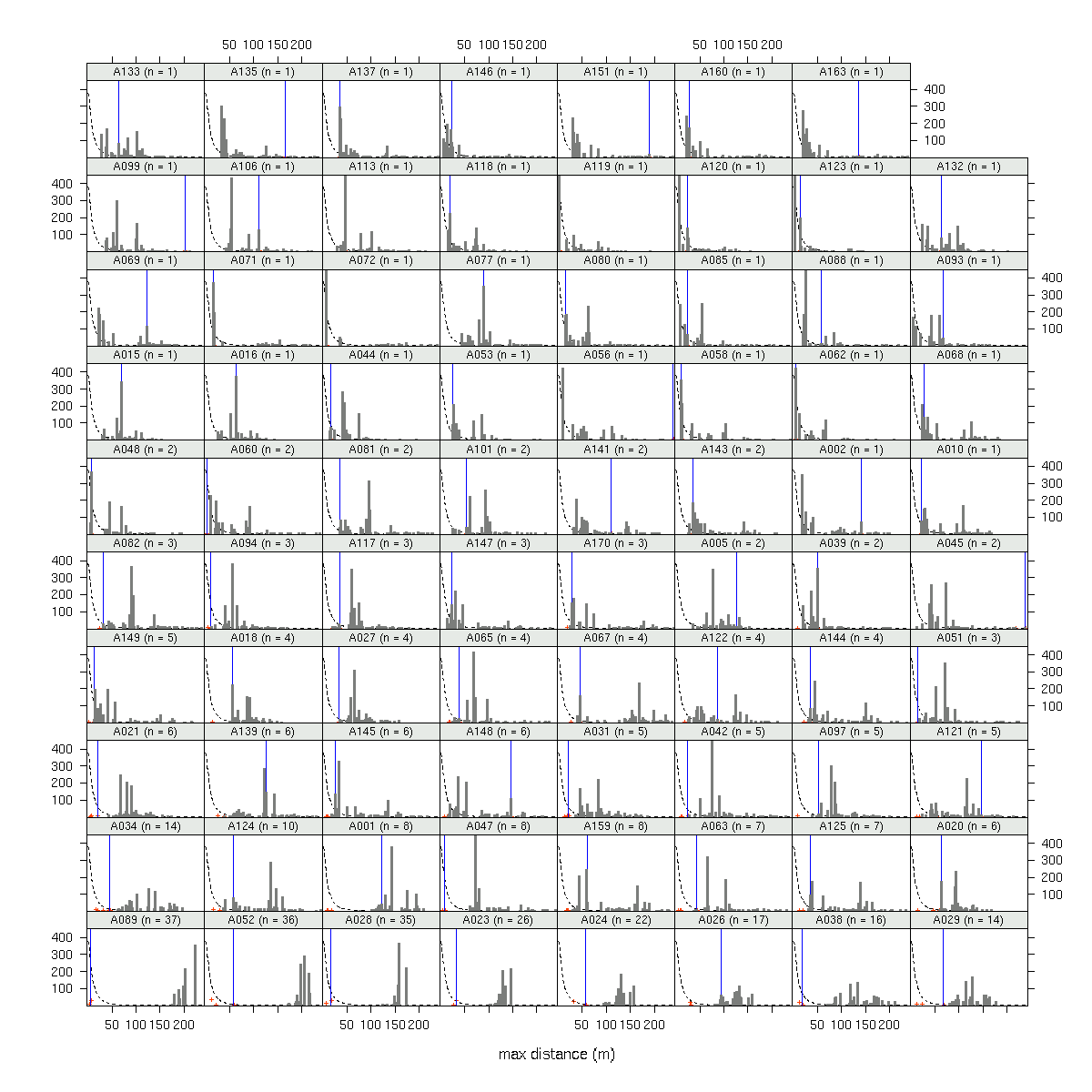
A089
(発見された子供数 37)
の場合.
fat-tail な散布カーネルを仮定した場合,
「最遠方に飛ばされた子供までの距離」
は 200m 超になる確率が高い
(しかし現実は 20m ぐらいだ)
ということがわかる.
つまり,
このぎょーかいでよく使われてる
「いつでも fat-tail 散布カーネルで OK!」
てなふうに使うのはダメ,
ということ
(観測されたパターンを説明できない).
A089
も含む全 79 母樹の library(lattice)
作図
(→ 全体).
先日の
母樹からみた倒木分布
という図も参考になるでしょう.

CH2O
でその分子量は 30?

beta2 ~ dnorm(beta2, tau)
という書きかたは許される,
とわかった.
これは randomwalk sampler になっているのだろうか?
……
と思ったんだけど,
考えてみるとわけのわからぬ尤度評価がなされているので,
使わない
ほうがよいワザ,
という気がする.
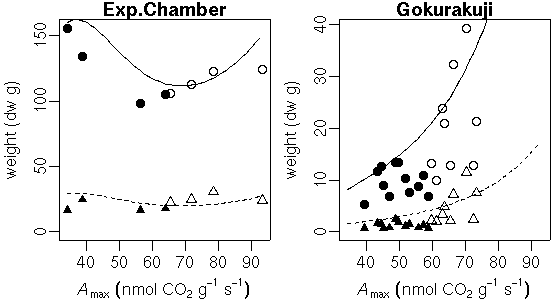
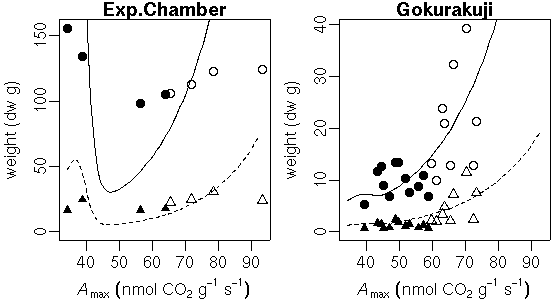
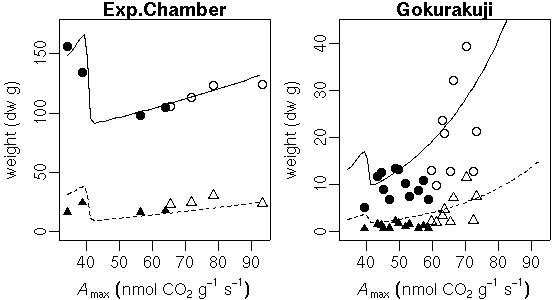
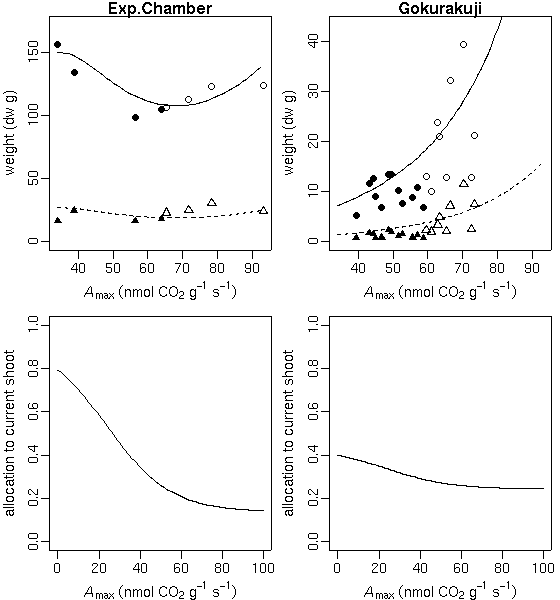
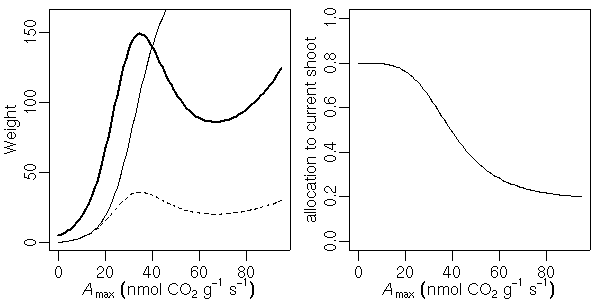
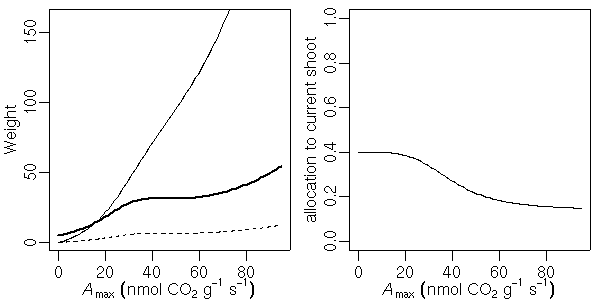
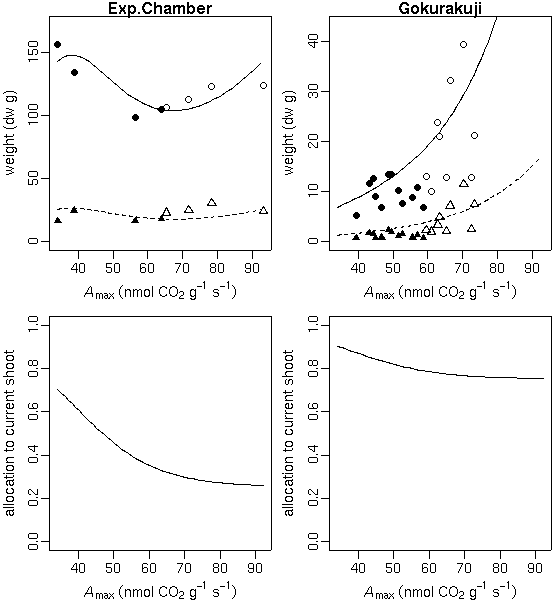
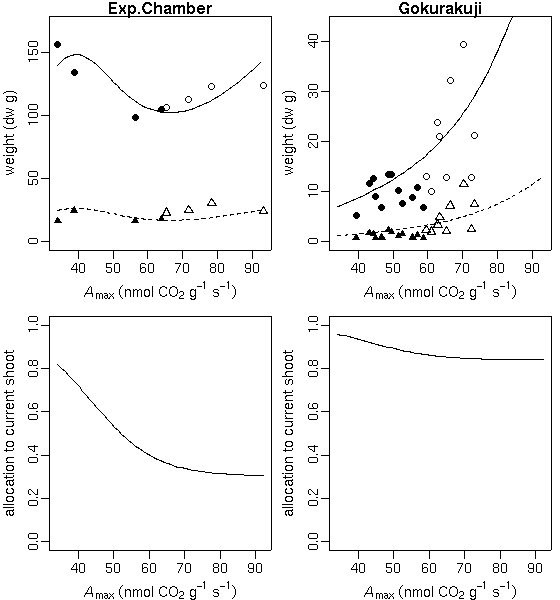
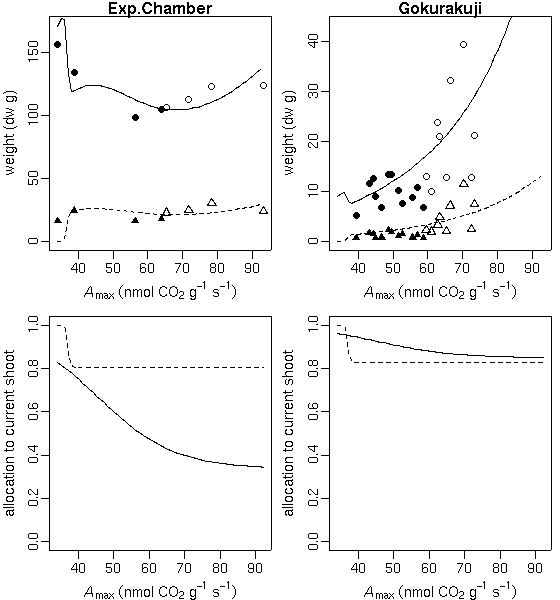
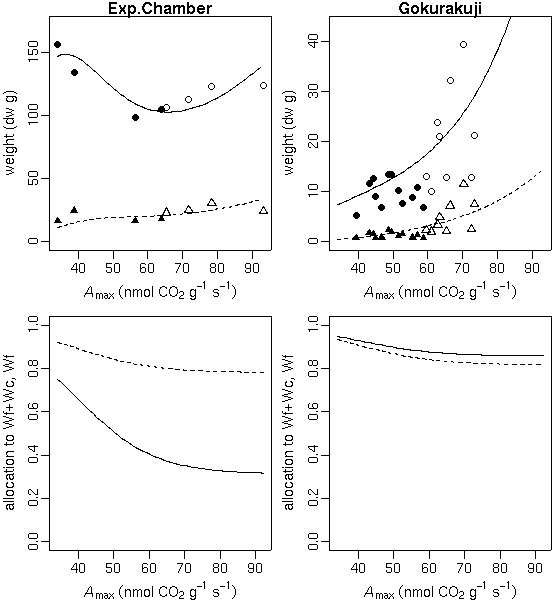
…… と同時に, 生理生態の分野で, こういう観測データが得られたときに, 成長段階の各時刻における資源分割をこうだと仮定した (素過程の組みあわせにすぎない) 簡単なシュート成長モデルを構築して, 観測データにあわせてそのパラ メーターを推定しました, というごく基本的なハナシなんかは 意外と考えられてないような気もします. 皆さん, 割算値に依存しすぎではないでしょうか ……
(abstract) We studied mechanisms of vegetation change in fens subject
to succession from open water to floating mats and finally herbaceous
rich-fens. Earlier research showed that these systems are characterized by
transient seed banks. Our main question was whether seedlings of later
succssional fen stages are already present in earlier stages, remaining
subordinate in the vegetation until conditions become suitable for them.
If, however, conditions during succession change in a way that only a
limited set of species can survive as seedlings during each of the succssional
stages, no seedling bank will exist. The transient character of the
seeds would then imply that seeds will not germinate and will subsequently
die and that seeds that have germinated in the ``wrong'' stage will not become
established. We hypothesized that: (1) germination and seedling survival
of fen species are significantly better in the succssional fen stage for which
these sepecies area characteristics. (2) as a consequence no seedling banks
occurs in these fens.
...
Germination was higher in the ``own'' stage for all species groups as was
survival for quaking fen species and rich fen species.
...
Because of the transient seed bank and absence of a seedling bank in these
fen wetlands, successful establishment of species necessitates a continuous
dispersal of characteristic species until the environmental conditions permit
establishment.
This also implies that species of the whole successional sere should be present
within dispersal distance.
(abstract) Many glassland restoration projects on former arable land face
problems because early successional grassland species establish vigorously
and persistently but late successional grassland species fail to establish.
Differences in establishment characteristics of early and late successional
species might provide an explanation for the failure of many late successional
species to colonize grasslands on ex-arable land.
...
The poor establishment of late successional species on former arable land
could therefore not be explained by solely by differences in establishment
characteristics between early and late successional grassland species.
Competitive processes between early and late successional species later in the
life cycle probably play an important role. The results do point out that
establishment of late successional species can be promoted by creating
vegetative cover from the start of the restoration effort.
(abstract) Questions: How do physical microsite conditions of microsites affect
germination and seedling survival in different succssional stages?
Do different species germinate in similar microsites in a given successional
stage?
...
Results: In early successional sites, seedlings of several species were
positively associated with depressions and presence of rocks, and negatively
associated with ridges. Patterns were generally consistent among species.
In later succession, seedlings were not signficantly associated with any
microsite characteristics. For Alnus viridis, seed density decreased
with distance from seed sources but seedling density did not.
Conclusions: Because of harsh conditions in early succession, physical
microsites are important, and most species have similar microsite requirements.
In later succession, physical microsites characteristics are not as important
and are more variable. Microsites appear to be more important than seed
rain in controlling the distribution of Alnus viridis in early succession.
(summary) 1. Changes in farming practice provide an opportunity to restore
once extensive forested wetlands on agricultural land. In some parts of the world,
however, it has proved difficult to restore the full complement of plant species
through natural regeneration. Similarly, the restoration of forested wetlands
by replanting has often resulted in ecosystems of low diversity. Better methods
of restoring these important ecosystems are now required and baldcypress swamps
provide and opportunity to investigate alternative approaches to the restoration
of forested wetlands. This study examined the composition of of seed banks of
farmed fields to determine their value in restoring swamps in the south-eastern
United States.
...
5. Synthesis and applications. Both restoration ecologists and managers
of nature conservation areas need to be cognisant of seed bank and dispersal
characteristics of species ot effectively restore and manage forested wetlands
In the case of baldcypress swamps, critical components of the vegetation are not
maintained in seed banks, which may make these floodplain wetlands difficult to
restore via natural recolonization. Ultimately, the successful restoration of
abandoned farm fields to forested wetlands may depend on the re-engineerring of
flood pulsing across landscapes to reconnect dispersal pathways.
(summary) 1. As an aid for restoration projects, the relative potential for
plants to immigrate to sites after severe disturbance was investigated and an
index developed to determine their relative immigration potential. The goal
was to establish baseline knowledge for the species pool in severely disturbed
sites prior to restoration. The method was based on the identification of
potential colonists, inventories of populations in surroundings vegetation
and autecological information on their maximum fecundity and dispersal by wind,
water or animals.
...
5. Synthesis and applications: An index of immigration potential provides
information on the probable initial species pool after severe disturbance,
against which the effects of local habitat suitability and establishment factors
can be added to assess probable successional patterns. Its flexibility and
applicability to varied life forms should make it useful for diagonosing
recolonization bottlenecks in a wide variety of disturbance and restoration
situations. It also allows for and assessment of the relative need for
introducing species during the restoration of severely disturbed environments.
(abstract) Question: Do species from communities with different flooding
dynamics differ in seed buoyancy? Is there a trade-off between seed buoyancy
and seed longevity?
...
Results: Community occurenence along the hydrologic gradient was mirrored by
seed buoyancy. Seed buoyancy was highest for speices of almost permanentaly
inundated reed beds and lowest for species for species of rarely
inundated wet meadows. The seed buoyancy characteristics of species
of reed beds were best adapted to hydrological dynamic conditions
mimicked by the treatment moving water and were the most independent of the
flow rate of the water. A high percentage of the species used had a transient
seed bank. Together with high in many species, this suggests a trade-off between
dispersal capacity and seed persistence.
Conclusions: In freshwater wetlands with rather stable water levels, many
species lack a persistent seed bank and depend on dispersal for colonization
of new habitats. Seed buoyancy enhances aquatic seed dispersal, and can be of
great importance in both vegetation dynamics and restoration. In wetlands
that are inundated for a long period during the year and with a water level
above soil surface, higher seed buoyancy enhances the possibility for
hydrochory.
(abstract) Seed production, composition of seed rain, germination and seedling
mortality, as well as vegetative growth characteristics of common pioneer
plant species were studied on the foreland of the retreating Morteratsch glacier
in the Swiss Alps. The frequency of diaspores trapped in different successional
stages was related to their dispersal mode and was highly skewed towards a few
species. Plenty of diaspores well adapted for dispersal by wind area a
precondition for the most important pioneer species. Seed from all pioneer
species investigated had a good germination success, provided that the moisture
content of the soil was high enough. However, requirement for seedling
establishment differed among sites of increasing terrain age and among species.
...
The life cycle of this species is characterized in succession by
(1) the colonization of safe sites by small seeds adapted for wind dispersal,
(2) horizontal spread by clonal growth, and (3) the persisitence through
phenotypic morphorogical plasticity and clonal growth are thus complementary
mechanishms in plant succession on recently deglaciated terrian.
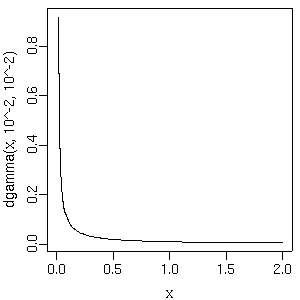
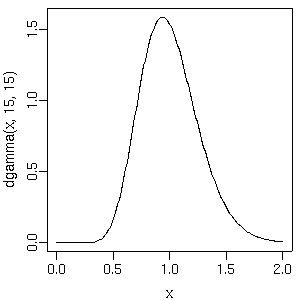
dlnorm()
(対数正規分布) にするとコケることがあるな
(slice sampler が動かないとかで).
回避わざとしては
(ばかばかしいけど)
log.beta2[j] ~ dnorm(0.0, Tau.noninformative) beta2[j] <- exp(log.beta2[j]) # もしくは log(beta2[j]) <- log.beta2[j]なぜかこれだと問題おこらない. どうもこの
dlnorm()
だの
dgamma()
だのといった「正の値しかとらない確率分布」
からの sampling は得意ではないみたいだねえ.

James S. Clark, Mike Dietze, Sukhendu Chakraborty, Pankaj K. Agarwal, Ines Ibanez, Shannon LaDeau, Mike Wolosin.
2007.
Resolving the biodiversity paradox.
Ecology Letters (OnlineEarly Articles).
doi:10.1111/j.1461-0248.2007.01041.x
いやはや,
この御大的 abstract って
……
The paradox of biodiversity involves three elements,
(i) mathematical models predict that species must differ in specific ways
in order to coexist as stable ecological communities,
(ii) such differences are difficult to identify,
yet (iii) there is widespread evidence of stability in natural communities.
Debate has centered on two views. The first explanation involves tradeoffs
along a small number of axes, including `colonization-competition',
resource competition (light, water, nitrogen for plants, including
`successional niche'), and life history (e.g. high-light growth vs. low-light
survival and few large vs. many small seeds).
The second view is neutrality, which assumes that species differences do not
contribute to dynamics.
Clark et al. (2004) presented a third explanations,
that coexistence is inherently high dimensional, but still depends on species
differences.
We demonstrte that neither traditional low-dimensional tradeoffs nor
neutrality can resolve the biodiversity paradox, in part by showing that
they do not property interpret stochasticity in statistical and theoretical
models. Unless sample size are small, traditional data modelling assures that
species will appear different in a few dimensions, but those differences will
rarely predict coexistence when parameter estimates are plugged into
theoretical models.
Contrary to standard interpretations, neutral models do not imply
...
う,
おもしろすぎてつい長々と写経してしまった.
ここで紹介されてる森林動態の階層ベイズの詳細は
Clark et al. (2004)
に書かれている.
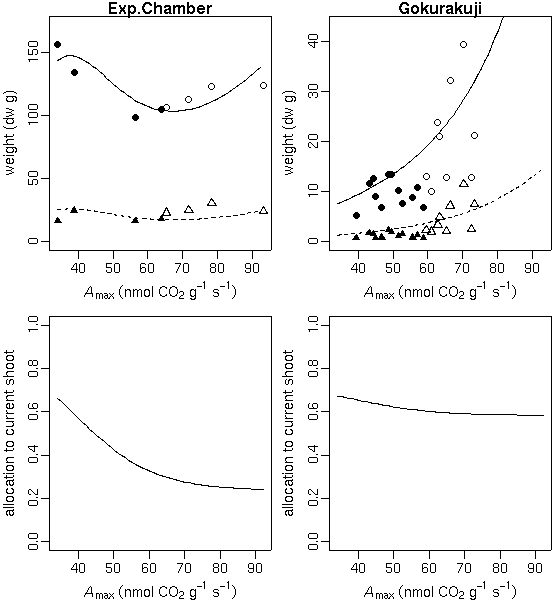
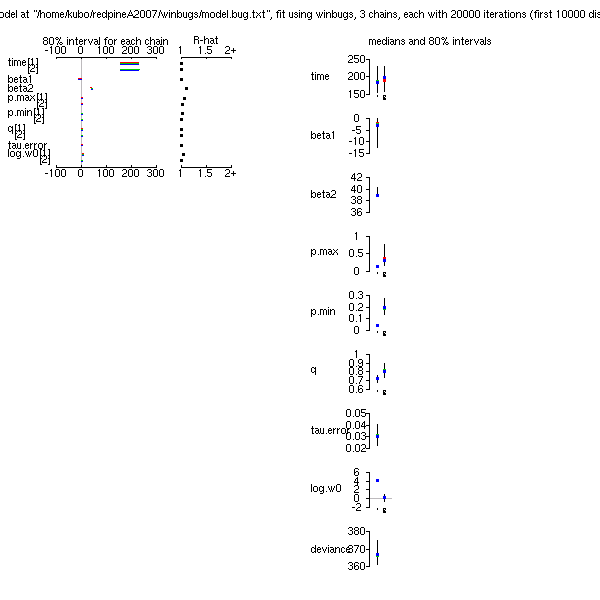
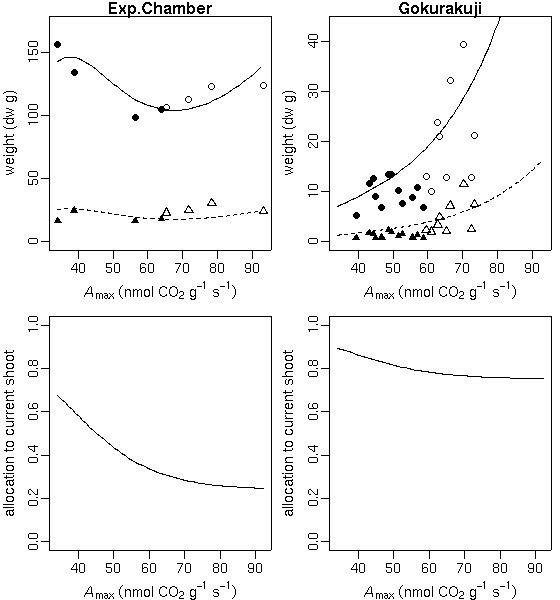
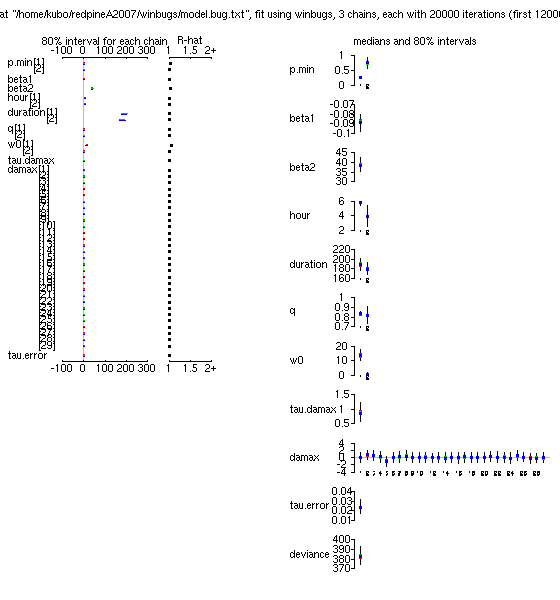
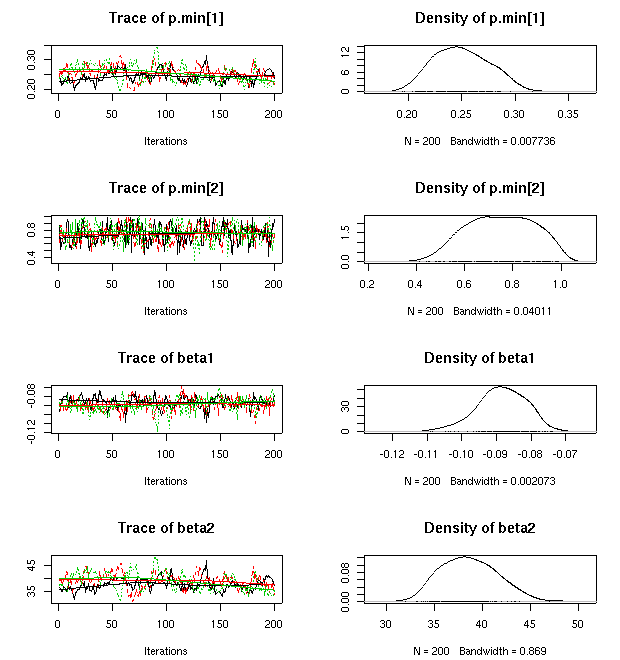
p.max
ってもはや不要なパラメーターだよな」
といまさらながら気づいたので,
これを削除してみる
……
やはり,
推定された「シーズン終わりの当年生葉重・茎重」予測はあまり変わらず.
x
ってのが「分散の逆数」
tau.damax
ね.
tau.damax
のとりうる範囲をこのように制約してやると,
「ずれ」
damax
(「個体差」)
もゼロからあまり逸脱しなくなるので,
まあ収束します,
と.
\newcounter{aff}
\renewcommand{\theaff}{\fnsymbol{aff}}
\newcommand{\Aff}{$^{\theaff}$\addtocounter{aff}{1}\ }
とプリアンブルで定義しておいて,
このように原稿を書いて,
\setcounter{aff}{1}
CHUNLAN LIAN,\Aff
SUSUMU GOTO,\Aff
TAKUYA KUBO,\Aff
\setcounter{aff}{2}
YASUO TAKAHASHI,\Aff
\setcounter{aff}{4}
MASAHIKO NAKAGAWA\Aff
and TAIZO HOGETSU\Aff
\begin{list}{}{\baselineskip=1mm\parskip=2mm\usecounter{aff}}
\renewcommand{\makelabel}{\theaff}
\item Asian Natural Environmental Science Center, the University of Tokyo,
Midori-cho 1-1-8, Nishitokyo-shi, Tokyo 188-0002, Japan.
\item University Forests in Hokkaido, the University of Tokyo,
Yamabe, Furano, Hokkaido 079-1561, Japan
\item Graduate School of Environmental Earth Science,
Hokkaido University, Sapporo, Hokkaido 060-0810, Japan
\item Hokkaido Forestry Research Institute, Koshunai, Bibai,
Hokkaido, 079-0198
\item Graduate School of Agricultural and Life Sciences, the
University of Tokyo, Yayoi 1-1-1, Bunkyo-ku, Tokyo 113-8657, Japan.
\end{list}
これをコンパイルするとこうなります,
と.

\subsection{} の書式の変えかたがわからん
……
と検索してみたらやはりぎょーむ日誌にのってた.
2001-08-09.
Molecular Ecology はこんなところかしらん?
\renewcommand{\section}{\@startsection
{section}{1}{\z@}
{-3.25ex \@plus -1ex \@minus -.2ex}
{1.5ex \@plus .2ex}
{\raggedright\normalfont\large\bf}
}
\renewcommand{\subsection}{\@startsection
{subsection}{2}{\z@}
{-3.25ex \@plus -1ex \@minus -.2ex}
{1.5ex \@plus .2ex}
{\raggedright\normalfont\itshape}
}
\makeatother
lv
で見てみたらなぜか日本語文字 (2 byte)
バックスラッシュつかった (化け) LaTeX コードだった
……
ってことでこれはゐんどーづ環境固有の問題なのか?
{{...}}
なるいんちきわざで bst formatting が回避されていた
(ので {...} に置換した).
ちゃんと書かれていれば問題ないんだけど,
提供もとのぢゃーなるによってまちまちで.
学名も斜体になってないし.
390 → 381
とせこく改善されている.
これは採択.
{kind=link}
{kind=link}
{kind=link}