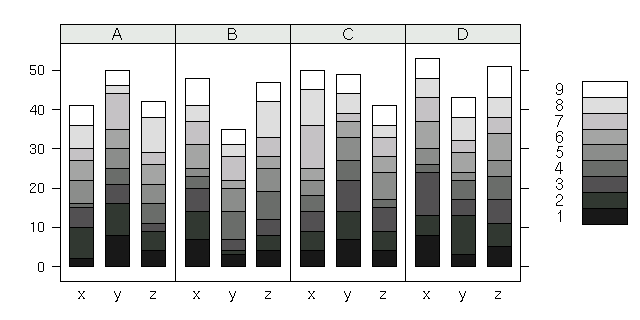ぎょーむ日誌 2007-07-(01-10)
2007 年 07 月 01 日 (日)
-
0800 起床.
コーヒー.
朝飯.
洗濯.
怠業
……
ばかりもしてられないので,
少しばかり部屋の掃除・かたづけ
……
で午後になってしまった.
昼飯.
-
まあいつまでもうだうだしてられないので,
1350 自宅発.
快晴かつすずしく札幌らしい初夏.
1405 研究室着.
A 棟 8F 無人状態でおちついているので,
まずはのんびりとお茶部屋でちょっと掃除 & かたづけ.
-
で,
さっさと仕事やればいいのに
……
こんどは KuboWeb 内のディレクトリ大移動.
といってもアクセスしてるヒトには何がどうなったかわからないだろうけど.
またしても Apache Web server の「黒魔術」
こと mod_rewrite
にめんどうを押しつけることに.
-
ぢりぢりとまたアリ作文.
やはり,
他人に原稿 (まだ図表とモデル説明と結果しか書いてないけど)
みてもらおうと議論要約などつくったりしていると,
ここまで書いた部分の不備 (用語の不統一など)
がぽろぽろと見つかるので,
その修正.
-
2020 研究室発.
2040 帰宅.
晩飯の準備.
晩飯.
-
[今日の運動]
-
腹筋運動 30 ×
3 回.
腕立ふせ 10 ×
3 回.
スクワット 100 回.
-
[今日の食卓]
- 朝 (0840):
イソップベイカリーのバタール.
ピーマン・ネギ・イカ・卵の炒めもの.
- 昼 (1250):
イソップベイカリーのバタール.
コマツナのマヨネイズサラダ.
手作りマヨネイズは難しい (酢の選択が悪かった?).
トマト.
ヨーグルト.
- 晩 (2230):
米麦 0.8 合.
ネギ・豆腐の味噌汁.
カツオのたたき +
タマネギスライス.
イカ・セロリの煮物.
ワカメ・キュウリ・トマトのサラダ.
2007 年 07 月 02 日 (月)
-
0800 起床.
コーヒー.
朝飯.
0920 自宅発.
晴.
0940 研究室着.
-
また
エゾアカヤマアリ (大統領命令) 拉致られアリいじめられ実験データ解析の
作文修正
……
も終わってきたので PDF 化 + ファイルアップロードしくみを整え,
議論概要かいた web page からダウンロードできるようにしてみる.
大統領と藤原君に説明メイルかいて送信.
あとは先方からの反応まち.
-
まち時間のあいだ,
先週,
紺野さんから送っていただいた
十勝平野の樹木繁殖データにちょっととりくんでみる.
1130 ごろから佐藤さん修論読んでみる
……
いやー,
さすがに紺野さん御指導だとたいへん丁寧な内容,
といいますか.
-
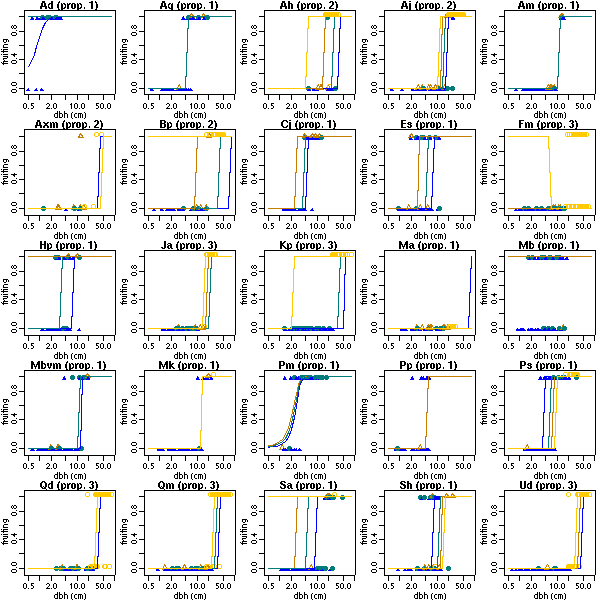
送っていただいたデータセットもさっそく拝見.
うーむ,
種数も個体数も多い
……
> summary(d$SP)
Ad Ag Ah Aj Am Axm Bp Cj Es Fm Hp Ja Kp
24 47 34 64 61 16 60 39 55 104 39 42 96
Ma Mb Mbvm Mk Pm Pp Ps Qd Qm Sa Sh Ud
45 64 50 50 70 37 90 108 124 65 56 92
とりあえず作図.
データ解析はつねに「とりあえず作図」から始まる.
-
これだけたくさんデータがあっても
「樹種ごとに glm()」
ってのはぜんぜんダメだとわかった.
それはいい.
このデータ解析はべいづでケリをつけるために私のところにまわってきたんだから.
しかし
……
やはり,
というかとうぜんながら
樹木サイズと「明るさ」の交絡がある
……
うーむ.
-
あららいつのまにか時刻はすでに 1345.
昼飯調達の旅にでよう
……
と思ったらアリ原稿について藤原君からメイル連絡.
岩倉さんの連絡先もわかったのでメイルしたためて送信.
1400 研究室発.
北大生協までふらふらと.
研究室にもどって昼飯.
-
で,
階層ベイズモデル化.
このモデリング,
まじめに考えると多々難しいところがあるんだけど,
いきなりややこしい (つまり実戦的な) 統計モデル化すると
紺野さん・佐藤さんにとってわけわからぬものになってしまう
……
ということで,
佐藤さん修論の logistic 回帰に最低限の拡張をくわえた
logistic な階層ベイズモデルを定義し,
その MCMC 計算を.
-
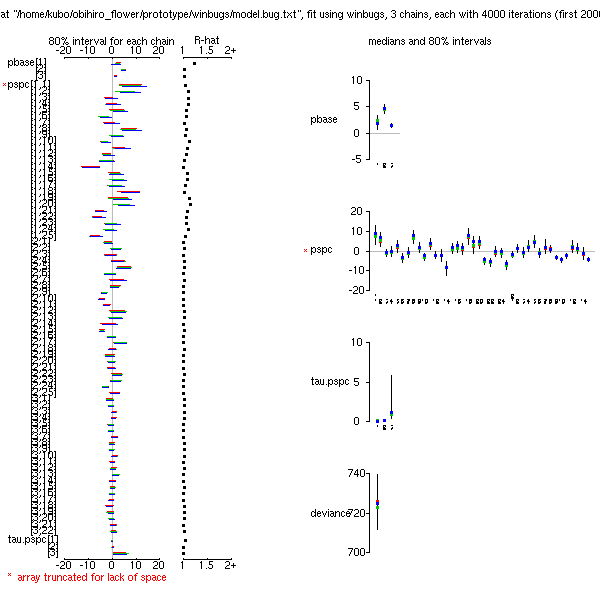
やはり収束が悪いな
……
ということで A801 院生部屋の Dell desktop Linux 機をたたきおこして,
高速計算もーどに移行する.
-
統計モデルがあまりよくない (?) せいか,
事後分布の収束があまりよくないのをチカラまかせに無理矢理計算.
4000 MCMC step トリプルチェインで 500 秒ちょい.
まあ,
こんなところでとりあえずは十分かな
(事後分布表).
-
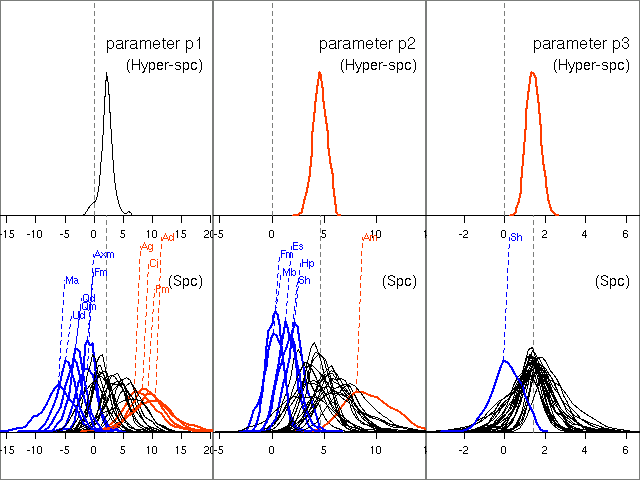
そして nested 事後分布図.
ほーら,
ね
……
こういう群集データでは
「種間共通部分」 vs 「種間相違部分」
のふたつを同時に考慮するベイズな手法がたいへんうまくいくのである.
-
ばててきたので撤退.
1950 研究室発.
2010 帰宅.
体重 67.8kg.
晩飯.
お,
アリの件に関して岩倉さんから連絡いただけた.
-
[今日の運動]
-
腹筋運動 30 ×
3 回.
腕立ふせ 10 ×
3 回.
スクワット 100 回.
-
[今日の食卓]
- 朝 (0830):
米麦 0.5 合.
ネギ・マイタケの味噌汁.
イカ・セロリの煮物.
ワカメ・キュウリ・トマトのサラダ.
コマツナのゴマあえ.
- 昼 (1430):
研究室お茶部屋.
北大生協のサツマイモリンゴパン.
トウモロコシ.
- 晩 (2040):
キャベツ・ニンジン・ネギ・セロリ・ショウガ・イカ・卵
焼きそば.
トマト.
コマツナのゴマあえ.
2007 年 07 月 03 日 (火)
-
0800 起床.
ねむい.
コーヒー.
朝飯.
0940 自宅発.
晴.
0955 研究室着.
-
Nature
vol. 447 最新号
ではサルの神経系は統計モデリング + パラメーター推定
(つまり確率論的な学習)
が可能かどうか調べた論文が.
さてさて,
石狩浜の「のほほんアリ」たちの神経系 (とくに触角の中の神経)
はどのように「学習」すると,
他のアリと接触したときにああいうふうに
(つまり階層ベイズモデルで解析した行動データみたいに)
応答しちゃうのか
……
というのを一ヶ月後までにあるていどは研究しないといけないんだよね.
やれやれ.
-
1030 より
研究室セミナー.
今日は斎藤君で真駒内とか放棄スキー場で「次」にでてくるのは
(在来な) ススキ群集なのか (外来な) オオアワダチソウ群集なのか,
そのあたり調べます,
というもの.
植生・発芽・食害・環境改変うんぬん,
といういかにも露崎弟子的な方向性なんだけど
……
うーむ,
野外実験っていろいろと難度たかそう.
ちなみに道内あちこちに放棄スキー場あるんだけど,
「次」になにが出てくるかは場所によって異なるそうで.
-
午後にとあるデータ解析こんさるが予定されてるんだけど,
難度の高い作図問題になりそうなので事前に研究をススめてみる.
やはり一時間ちょっとを費やして,
とりあえずはできた.
[barchart() による「組成」ぐらふ]
library(lattice)
の
barchart()
を使って
barchart(n ~ treatment | forest, groups = spc, data = de2, stack = TRUE, col = rainbow(9))
とすると左のような図になる.
データ格納してる data.frame である
de2
については下を参照,
めんどくさい
reshape()
つかって data.frame を
「長いカタチ」
(
direction = "long")
変換するところがぽいんとである.
> data.example # こういうデータがあったとする
forest treatment spc.1 spc.2 spc.3 spc.4 spc.5 spc.6 spc.7 spc.8 spc.9
1 A x 7 4 3 3 5 2 4 6 6
2 B x 3 9 5 5 4 1 3 5 5
3 C x 2 5 7 4 6 2 7 5 2
4 D x 4 5 5 2 4 6 5 9 5
5 A y 9 7 9 7 12 5 4 8 4
6 B y 2 5 5 3 4 2 1 6 4
7 C y 3 3 4 7 4 6 4 7 8
8 D y 4 3 3 9 8 4 3 5 4
9 A z 5 4 4 2 3 7 4 9 5
10 B z 4 4 7 5 4 6 4 7 6
11 C z 0 8 7 5 3 5 4 4 7
12 D z 5 3 5 7 4 7 5 5 1
> v.spc # こういう列名 vector をあらかじめ準備しとく
[1] "spc.1" "spc.2" "spc.3" "spc.4" "spc.5" "spc.6" "spc.7" "spc.8" "spc.9"
> de2 <- reshape(data.example, direction = "long", varying = v.spc)
> de2 <- de2[, 1:4]
> colnames(de2) <- c("forest", "treatment", "spc", "n")
> de2 # reshape() によって「長いカタチ」に変換された data.example
forest treatment spc n
1.1 A x 1 7
2.1 B x 1 3
3.1 C x 1 2
4.1 D x 1 4
5.1 A y 1 9
6.1 B y 1 2
7.1 C y 1 3
8.1 D y 1 4
9.1 A z 1 5
10.1 B z 1 4
11.1 C z 1 0
12.1 D z 1 5
1.2 A x 2 4
2.2 B x 2 9
3.2 C x 2 5
4.2 D x 2 5
5.2 A y 2 7
... (以下略) ...
-
まあ,
library(lattice)
つかわぬ,
ふつーの
barplot()
でも,
つみあげな図は描けたりするわけですが
(RjpWiki 内の例).
-
とりあえず昼飯.
-
で,
1400 すぎに農学部の高橋さんがみえて,
R 作図難問の検討.
上述のごとく
library(lattice)
の
barchart()
で正解なんだけど,
さらに
library(lattice) マニュアル
の研究が必要で
-
ハコ塗りの彩色をユーザー指定にするための
trellis.par.get("superpose.polygon")
設定わざ
-
凡例だすための
auto.key = list(...)
わざ
-
さらに凡例を上下逆転させる
reverse.rows = TRUE
指定
(詳細は久保作関数の
barchart.gray.R
参照してください)
といったあたりが難所だった.
たとえば上と同じデータ構造をもつ
de2
(ただし乱数でつくったニセデータの数値は上とは異なる)
の表示はこうなる.
> source("barchart.gray.R") # 関数定義のファイル名が "barchart.gray.R" の場合
> barchart.gray(de2, v.spc) # de2 だの v.spc だのは上を参照
-
どーにかこーにか作図難問が終了した
……
と,
お茶部屋で脱力してるとまたここで西村さんと
「被度」解析なデータ解析検討あれこれ
……
「被度」はやっぱりめんどうですなぁ.
データ解析者からすると,
「目で見てハカる被度」より
10 × 10
コぐらいに切ったワクつかった
「ある・なし」データな「被度」のほうがマシ,
というのも,
これで二項分布つかった統計モデリングに正当化の根拠をあたえるから,
です.
-
1955 研究室発.
2010 帰宅.
晩飯の準備.
晩飯.
-
2220 自宅発北大構内走.
2310 帰宅.
-
[今日の運動]
-
[今日の食卓]
- 朝 (0840):
キャベツ・ニンジン・ネギ・セロリ・ショウガ・イカ・卵
焼きそば.
- 昼 (1340):
研究室お茶部屋.
食パン.
- 晩 (2110):
米麦 0.7 合.
ワカメ・豆腐のスープ.
キュウリ・トマト.
2007 年 07 月 04 日 (水)
-
0700 起床.
コーヒー.
朝飯.
0825 自宅発.
晴.
0840 研究室着.
-
とりえあえずエゾアカヤマアリに関して,
大統領に談判もうしこむメイル書いてみる.
ほっとくと放置されるだろうからなあ
……
-
このメイルもしばらく放置されるだろうから,
その間に一昨日の帯広樹木繁殖の (説明用に簡単化した)
階層ベイズモデルの説明かき.
なんだかんだいっても
VikiWiki
はお手軽だなぁ
(左上の図は佐藤さん修論 ppt ファイルから借用).
-
1030 すぎに紺野さんにメイル送信してひと区切り,
かしらん?
とりあえず
(意外にも放置されずに)
大統領談判メイルに応答あったので,
拉致られアリまわりでも再整備するか
……
-
で,
アリ論文ながめてて気づいたんだけど,
来月アタマの沖縄行脚の旅行手配をまったくやってない!
とりあえず航空券.
えーと,
8/3 (金) 発の 8/9 (木) もどる,
か.
この時期の沖縄旅行者のうちもっとも楽しくない気分の一人,
となるんだろうな.
-
旅程とかうだうだ考えながらだとなかなか時間かかる
……
航空券は帰路がややこみぎみ,
宿泊はぜんぜんこんでないかんじ.
航空運賃 (往復) 88300 円,
宿泊 (やんばる抑留期間除く,那覇市内の三泊朝食つき)
17610 円.
うーむ.
そして時刻はすでに正午すぎ.
-
ハードウェア買いの好きな甲山さんから
A804 セミナー室用の無線 LAN るーたー買い命令が
……
来月のなんちゃら講習に使うそーで.
-
昼飯.
アリ談判連絡は途絶中.
-
アリ論文よみ.
読んでるうちにふと今さらながら気づいたんだけど,
エゾアカヤマアリとアルゼンチンアリにみょーな符合があるような気がしてきた.
-
エゾアカヤマアリは定山渓では monodomous で
他巣のアリにたいして攻撃的である
:
アルゼンチンにおけるアルゼンチンアリも同様,
と言われている
-
石狩浜ではエゾアカヤマリは
すーぱーころにーとやらを形成し,
他巣のアリに対してもまーまーいいぢゃないですか
とあまり敵対的ではない
:
アルゼンチンアリも侵入先
(地中海まわりだの北米だの日本だの)
では同様,
と言われている
(一部の)
アリなヒトたちは遺伝子決戦主義とでもいうべき,
遺伝的多様性の大小で
原産地 vs 侵入先におけるふるまいの違いを
ぜんぶ説明できる,
としてるみたいだけど
……
そういうことではないよーな気もする.
たとえば,
アリの敵対性認識は触角で感知しうる
体表面炭化水素組成とかのたぐいだろうけど,
この炭化水素組成なんてのは日々
(は大げさか?)
ころころ変わっていくらしいんで,
はたして遺伝子がそれにどれだけ関与してるのやら,
という一点からみてもアヤしい,
と.
-
まあ,
アリでも (いわゆる) 「青ひげ」遺伝子らしきモノ,
ってのも見つかってるというハナシもあるけどね
……
-
六ヶ国協議における中心的に位置いすわる某国のごときふるまい,
というか
……
アリ談判は来週に順延することにした今日はさよならメイルいただいたので,
腹いせにアリ原稿 PDF ファイルつきメイルを返信する
(私はふだんはゴミつきメイルは送信しないようにしてるんだけど).
-
アリぎょーむはほうりだして,
地衣類繁殖の階層ベイズモデル論文の作図宿題をかたづけることに.
いつものごとく
R
による作図のプログラミング
……
こんなものかな?
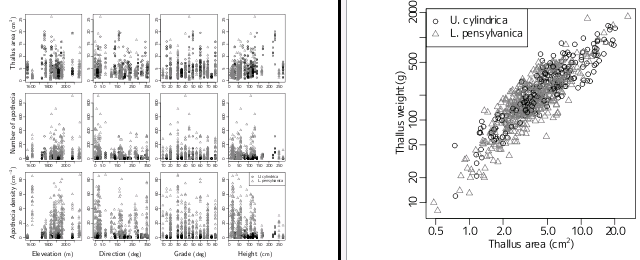
-
おっと,
そうだ,
予測された apothecia 数も図示する,
というハナシだったな
……
これはなかなかめんどう.
とりあえず中央値表示.
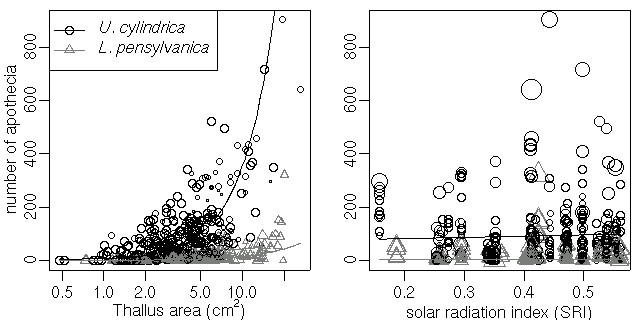
-
ともあれ (こればっかりは R + LaTeX でまとめている)
図一式 PDF ファイルをアップロードして志水さんにメイルかき.
-
紺野さんから樹木繁殖モデルに関するメイルいただいたので,
返信かき.
-
フィールドセンターの Nina さんから WinBUGS に関する
(まあ私にとって自業自得な) 難問メイルが.
英語にも堪能な
Nina さんに「おてがる生態学べいづ本」
ということで McCarthy のカエル本こと
Bayesian Methods for Ecology
(2007) お貸ししたんだけど,
その
例題 code
がどうも動いてくれない,
とのこと.
-
そして,
何やらうまくいかないトコロってのを
当方で再現しようとしたんだけど
……
えーい,
この例題とやらはゐんばぐすのアタマいたくなる user interface
の直接操作を要求するものなのである!
library(R2WinBUGS)
による遠隔操作ばっかりやってきた私には
……
-
幸か不幸か,
ゐんばぐす開発者たちもこのインターフェイスがいささか苦しいモノである
(とゆーか,
階層ベイズモデルを GUI で操作しよう,
という発想じたいが英雄的にすぎるのである),
とよく自覚してるためか,
WinBUGS project サイト内に
`` WinBUGS - the movie''
なるぽこぽこ flash file があり,
これを見れば誰でもゐんばぐす GUI 使いになれる
というありがたいモノがあった.
-
で,
Linux 上の
Wine
経由で WinBUGS 動かして
(この場合 Wine と X11 のあいだにけっこう深刻なる相性問題が存在する,
とわかった),
上記 McCarthy カエル本サイトからダウンロードした例題を
いくつか実行してみたんだが
……
何の問題もなく事後分布が得られた
(しかしこのボタンおしまくりのゐんばぐす GUI では研究にならづ,
やはり R2WinBUGS
が必要不可欠と確認できた次第).
-
ということで,
Nina さんに「どうしてこちらでは計算できちゃったのでしょう,
私には理由がわかりません,たすけてー」
と救援メイルに救援メイルで返してしまって一件落着
……
じゃなくてこの件,
先方の調査まち,
ということで.
-
おなかもへってきたし,
これ以上ややこしくならぬうちに逃げよう
……
ということで
1900 研究室発.
1915 帰宅.
晩飯の準備.
晩飯.
-
[今日の運動]
-
[今日の食卓]
- 朝 (0730):
米麦 0.6 合.
ワカメ・豆腐のスープ.
- 昼 (1320):
研究室お茶部屋.
食パン.
- 晩 (2050):
米麦 0.8 合.
ジャガイモ・ニンジン・タマネギ・トマトのカレー.
2007 年 07 月 05 日 (木)
-
0640 起床.
ねむい.
コーヒー.
朝飯.
0825 自宅発.
曇.
0840 研究室着.
アリ文献とかながめる.
-
0930 より教授会.
いつものごとくせれもにー的な.
しかし,
この時間はネットから隔離されるので文献よみに集中できて良い
……
ハナシも片耳では聞いてますよ.
この大学院だけの規模のそれほど大きくない
研究院の予算規模がいっさいがっさいで二億円ちょいといったケタ.
そんなに使うものなのね,
とちょっと驚いた.
助教の教員研究経費とやらは 52.8 万円で
助手のときの倍額だそーで.
もちろんこのおカネは何か研究室内の日々の消耗品とかで
「知らないうちに」
というかんじで消えていってしまう
(べつにそれで不都合ないけど).
-
最後に何やら来年の洞爺湖サミットに便乗して露出してみたい
北大の企てにすごく興味もつせんせーがたがあれこれ述べて
(景気よくなったせいで新入院生減ったあたりにも危機感おぼえているらしい),
1100 ごろ終了.
長びいた理由のひとつは,
ひさしぶりに「蛇行する」甲山さんとーくが二回ほど発動したため,
かしらん?
-
志水さんが研究室にきていたので,
昨日つくった地衣類論文の図について相談.
いくつか作りなおし.
サーヴァーにアップロード.
-
1215 研究室発.
家にもどって昼飯たべる.
ついでに洗濯.
1320 研究室もどる.
-
東さん秘書の三浦さんから沖縄出張に関する連絡いただいたんだが
……
かなり腐れている (JTB 製?)
北海道大学旅費システム
に苦闘させられる.
まず,
何がどうなってんのか,
Mozilla 系ぶらうざ (Galeon や Firefox)
ではログインすると frame だし無限ループにおちこむ.
Linux 上で動作する web browser のうち,
暴走しないのは Opera だけである.
しかも 9.20 みたいなのはダメで,
9.0* ぐらいぢゃないとログインできない.
-
で,
依頼出張の場合は「大統領から出張命令が」という
(旅費システムが自動発信する)
メイルうけとったら,
つぎにあたかも自分が旅行申請するかのようにしないといけないのね
……
で,「依頼出張」フォームに入り込む,
と
……
といったことを教えていただいた.
うう.
-
そして 8/4 (土) 午前の那覇→やんばる移動は大統領のきまぐれによって
現場で決定される
……
という何ともアブない状況らしい,
と判明した.
そこで安全のため,
私の 8/3 (金) 宿泊するホテルを予約変更することに.
ふーむ,
首里城ふもと
(?) のやや値段たかめのところ,
か……
-
またトナー雑用あれこれ.
-
アリ文献を
……
と思いつつもなぜか統計学文献の整理に没頭してしまった.
まあ,
「次」のアリ研究に役たちそうな neural network 論文とか見つかったし.
-
で,
neural network → 多変量解析 →
多次元尺度構成法 (MDS) → 非計量多次元尺度構成法 (NMDS)
……
と勉強の方向がどんどん逸脱していって,
ですね.
しかし,
おかげでというか,
この凶々しき世界における NMDS の位置みたいなものが
よーやくにしてわかるようになってきた
……
いやいや,
自分のデータ解析ではぜったいに使いたくないワザですけどね.
-
粕谷さんから来週のセミナーの要旨をいただいたので,
セミナー web page をつくり,
メイリングリストに連絡をだす.
jeconet の皆様
北大の久保です.7/12 (木) の Trendy seminar (こちらの研究室で主催してい
るセミナーです) で九州大学の粕谷英一さんに話していただけることになりま
した (粕谷さんは 7/11(水) からしばらく北大で滞在研究されます).
セミナー案内: http://hosho.ees.hokudai.ac.jp/~trendy/2007/abstract/trendy152.html
日時: 7/12 (木) 16:00 より
場所: 北大・環境科学院 A 棟 8F A804 室
http://hosho.ees.hokudai.ac.jp/~trendy/img/map.png
今回は「二本だて」セミナーで,
[1] 16:00 より「近親交配は有利か不利か」
(10 分ほど休憩)
[2] 17:10 より「ノンパラメトリックな検定の光と影」
を予定しています.どちらか一方だけの参加も可能です.どなたでも自由に
参加していただけますので,お気軽におこしください.
要旨:
−−−−−−
近親交配は有利か不利か
粕谷英一(九大・理・生物)
近親交配はサイズ依存的な交尾と並んでもっともよく調べられてきた非ランダムな
交配であり、交尾行動の進化だけでなく、生態学や集団遺伝学のさまざまな観点から
研究されてきた。一般には、近親交配は不利であり近親交配回避は有利であると見な
されることが多い。理論的には、近親交配回避が有利なのか不利なのかは、血縁度と
近交弱勢のバランスで決まる。近親交配が有利でも不利でもないような近交弱勢の閾
値は比較的高いところにあり、近交弱勢がそれよりも弱いときには、近親交配回避は
不利で近親交配した方が有利である。自然条件下では、交尾相手を選ぶうえでの血縁
関係の役割は意外なほどよくわかっていない。血縁個体が交尾相手として選ばれにく
いという報告もあり、一方で兄弟姉妹間のような近い血縁個体間での近親交配が普通
に見られる動物も少なくない。近親交配/近親交配回避を交尾相手を選ぶことである
とみたとき、これまで注目されていないどのような現象が見られると予測されるか述
べ、今までの少ない研究例との関係を検討する。
−−−−−−
ノンパラメトリックな検定の光と影
粕谷英一(九大・理・生物)
Mann-WhitneyのU検定、Kruskal-Wallisの検定、KendallやSpearmanの順位相関係数
、Jonckheereの傾向性仮説の検定などを代表的な例とするノンパラメトリックな検定
は、生物学に限らずデータ解析で広く使われてきた。データ解析に実際に携わる人々
の間での、その評価は、順位にするために情報をロスする鈍感な方法、頑健で使いや
すい方法、時代遅れな方法など、さまざまである。だが、ノンパラメトリックな検定
の実際の長所と弱点は意外に知られておらず、データ解析の実践家による妥当な評価
を困難にしている。正規線形モデルの方法(たとえば、t検定や分散分析)との比較
では、母集団の分布が何であってもt検定以上の検出力を持つノンパラメトリックな
検定が必ずあり、母集団が正規分布から外れればt検定を超える検出力を持つノンパ
ラメトリックな検定があることが昔から知られている。順位をとることでの情報のロ
スは多くの場合にはわずかである。一方、多くのノンパラメトリックな検定は等分散
を前提にしており不等分散の状況には不向きである。だが、データが顕著な不等分散
を示すときに好んでノンパラメトリックな検定を使用する、”自殺的”解析はしばし
ば見られる。生態学でのデータ解析を考えたとき、おそらくノンパラメトリックな検
定の最大の弱点は説明変数が複数のときにあり、とくに交互作用の扱いに顕著に表れる。
−−−−−−
1950 研究室発.
買いもの.
2010 帰宅.
体重 68.2kg.
あらら,
体重ちょっと増加ぎみだな.
晩飯の準備.
晩飯.
[今日の運動]
-
腹筋運動 30 ×
3 回.
腕立ふせ 10 ×
3 回.
スクワット 100 回.
-
ラジオ体操
(← 体操 PDF file あり:
そして自分で MP3 ファイルつくった).
ストレッチ.
[今日の食卓]
- 朝 (0700):
米麦 0.5 合.
ジャガイモ・ニンジン・タマネギ・トマトのカレー.
- 昼 (1240):
米麦 0.5 合.
ジャガイモ・ニンジン・タマネギ・トマトのカレー.
- 晩 (2230):
米麦 0.8 合.
ニンジン・ニラ・ネギ・ブナシメジ・ホタテの炒めもの.
ダイコン・トマト・レタス・ロースハムのサラダ.
コマツナのゴマあえ.
2007 年 07 月 06 日 (金)
-
0830 起床.
コーヒー.
朝飯.
0950 自宅発.
曇.
1005 研究室着.
-
ここまでのアリ作文みなおし & アリ文献よみ
……
に取り組もうとしてるんだけど,
なかななか進捗せぬまま昼飯.
-
``Polydomy in ants'' (2007, アリにおける多巣性)
なる最新の総説をみつけてしまった.
役にたちそう.
しかし,
なんともめんどうな
……
・ Introduction
- Interest of studying polydomy
- Definitions of polydomy
- Reminder of confusing terms
- Confusing situations
・ Polydomy as by-product of polygyny?
・ The syndrome of polydomy in ants
・ Ecology of polydomy
- Polydomy as a response to environmental heterogeneity
- Polydomy and the efficiency of resource capture
- Polydomy as a characteristic trait of tramp/invasive ant species
・ Consequences of polydomy for social evolution
- Polydomy and within-colony conflicts
- A new level of selection: the nest-level
- Modifications in reproductive and sex allocations
- Variability in colony closure
- Spatial distribution of nests within a population and its consequences
for genetic structure
・ A methodology to describe polydomy and to explore its effects
- Degree of colony closure
- Genetic analysis
- Determination of social structure
・ What remains to be discovered about polydomy in ants?
……
で一日がオワってしまいましたよ.
2125 研究室発.
2140 ばてばて帰宅.
晩飯.
[今日の運動]
[今日の食卓]
- 朝 (0900):
米麦 0.5 合.
ニンジン・ニラ・ネギ・ブナシメジ・ホタテの炒めもの.
ダイコン・トマト・レタス・ロースハムのサラダ.
コマツナのゴマあえ.
- 昼 (1310):
研究室お茶部屋.
食パン.
トマト.
- 晩 (2300):
米麦 0.7 合.
ダイコン・ニンジン・ニラ・ブナシメジ・干しエビのスープ.
キュウリ.
2007 年 07 月 07 日 (土)
-
0850 起床.
コーヒー.
朝飯.
怠業
……
というか,
ぼーっとしてた.
1030 ごろから洗濯.
-
怠業ばかりしてるわけにもいかんので,
昨日の Polydomy in ants 解読つづき.
VikiWiki 上でメモとりつつ.
-
1150 自宅発.
晴.
北大クラーク会館地下
理髪店
で散髪.
また 二ヶ月ぶり
になってしまった.
まち時間,
多巣アリ総説論文よみ.
1230 アタマをかりはじめて,
50 分ほどで終了.
1330 研究室着.
昼飯.

[変わっていく北大東側]
道路拡張とかで,
地環研すぐ前の
定食屋天勝だの,
その隣のよくわからなかった無線機屋だのがとり壊しとなった.

[北大内カラス繁殖地]
繁殖期のカラスはいらいらしてるから気をつけてね,
という注意書き.
百周年記念会館まわりで.
-
アリ総説よみ,
つづく.
-
1815 研究室発.
1830 帰宅.
晩飯の準備.
晩飯.
-
[今日の運動]
-
[今日の食卓]
- 朝 (0920):
食パン.
チーズ.
トマト.
キュウリ.
- 昼 (1400):
研究室お茶部屋.
食パン.
ワカメスープ.
- 晩 (1940):
米麦 0.7 合.
ジャガイモ・タマネギ・ソーセイジのスープ.
キュウリ・ニンジン・ブナピー・ささみ・春雨のサラダ.
2007 年 07 月 08 日 (日)
-
0900 起床.
コーヒー.
朝飯.
1055 自宅発.
快晴.
1115 研究室着.
-
岩倉さんからアリ拉致実験についていろいろとメイルで教えていただいた.
「たぶん,こうだろう」
という憶測で作ったモデルだけど,
「ほぼ正しい」
とわかった.
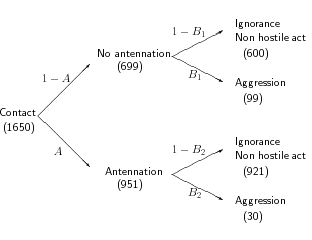
[行動連鎖の仮定]
まず antennation する・しないがあって,
次に攻撃する・しない,
と仮定しているわけだが
……
今さらながら,
この仮定が正しいかどうかは元データをあたればよい,
と気づいた.
で
grep
してみると
……
$ lv allinone.txt | grep " , a"
1 a a "b , a" a i jozankei jozankei30m o
-
……
また定山渓アリ
の凶行か.
全 1650 接触のうち
Jozankei vs. Jozankei の一ヶ所で
「攻撃 → antennation」
があるな.
この場合は地元アリが (30 m 離れた地点で拉致された)
拉致られアリに (瞬間的な判断で) 噛みついて,
さらに antennation しているのである.
「また定山渓」
というのは,
やはり全接触のうち一ヶ所だけ
「拉致られアリが地元アリ (Hoshioki) に逆襲」
という event があり,
その「逆襲した拉致られアリ」が定山渓出身なのである
(ぎょーむ日誌
2007-06-11).
-
まあ,
これはまたあとでデータ変換からやりなおして,
階層ベイズモデルの MCMC 計算もやりなおしておくことにしよう.
どうせここ一ヶ所かえても結果はほとんど変わらないし
(なにしろ Jozankei vs. Jozankei 状況における event だし),
R
によって
作図・作表は自動化
(投稿可能しあがり水準)
されてるので何べんで推定計算やりなおしもどうってことない.
-
といった返信メイルかいて昼飯.
-
で,
昼飯中に推定の再計算は終了
……
したと思ったけど,
やはり「30 m 離れたところで拉致」「Jozankei vs. Jozankei」
の交絡は依然として
「スキあらば収束悪化させる」
要因だとわかった.
-
で,
事前分布を工夫して,
いわゆる「切片」こと定数項パラメーターではない
fixed effects パラメーターは無情報ではなく
超事前分布に分散を制約される (IID) 事前分布をもつパラメーター,
としてみる
……
これで収束はうまくいき,
以前と同じ結果になった
(事後分布 summary table).
そして全ての図表の自動再生成,
と.
-
といった試行錯誤の計算まち時間にも多巣アリ総説論文をちらちらと読みつつ.
なんか,
アルゼンチンアリに関しては
(正しいのかどうかよくわからんけど)
遺伝的多様性でハナシをまとめようとしてるね.
だとしたら,
遺伝的なんちゃらでは
場所による挙動のちがい説明できそうにないエゾアカヤマアリはおもしろそう,
ということなのか?
-
1650 研究室発.
1710 帰宅.
晩飯の準備.
晩飯.
-
1925 より,
札幌駅すぐ北の小映画館
蠍座
で
「フラガール」
みた.
前回
みた「インディアン」をいかにもな「男の子」映画だとすると,
こちらは由緒正しい「女の子 (集団)」映画,
とするべきか.
好奇心旺盛で姿勢ただしく柔軟性と実行力にとみ理性的だけど
情理をわきまえ思いやりのある
「女の子」
たち (中には高齢の「女の子」も含まれる)
が果敢に問題にとりくみ,
悲しいことがあって泣き,
みんなで助けあい,
みんなどんどんキレイになっていって,
問題を解決して嬉しくて泣く映画
……
まあ,
今回もまた正調なるエンターテイメントです.
男どもはひきたて役というか道化役で,
立ちションして水かけられたり自分でハサんでしまったり,
ハワイ樹木ふぇちになってヤシに抱きついたり,
硬直した議論ふりまわして周囲を困惑させてみたり,
せいぜい借金とり相手にツルハシふりまわして借用書を食ってしまう,
ぐらいの役まわりで.
(後記)
書き忘れてたけど,
この映画みると平八郎提督の建白による
北海道ハワイ化計画
って実現するためにはそうとうの気合いが必要だとしみじみとわかった.
-
[今日の運動]
-
[今日の食卓]
- 朝 (0940):
イソップベイカリーの巨大クロワッサン.
- 昼 (1330):
研究室お茶部屋.
米麦 0.5 合.
フリーズドライ春雨スープ.
- 晩 (1840):
米麦 0.6 合.
麻婆豆腐,
ためしてガッテン
ふう.
キャベツ・ニンジン・タマネギ・キュウリ・レタスのサラダ.
2007 年 07 月 09 日 (月)
-
0800 起床.
コーヒー.
朝飯.
0925 自宅発.
曇.
0940 研究室着.
-
起床 → 登校の過程で,
アリ行動の階層ベイズモデルのハナシについて検討してたんだけど,
-
すーぱーころにーとやらでは同種内別巣アリへの
「nestmate 認識能力」
が欠落している,
というハナシがある
-
これは
「攻撃確率の低さ」
ってことでそう判断されている
-
エゾアカヤマアリに関していうと,
antennation では少なくとも二水準の強度,
攻撃に関しては三水準の強度があり,
「nestmate 認識能力」
がなくなっているわけではなさそうだけど
攻撃確率は全体に高くない
(1 接触あたりの確率がどのような場合でも 0.5
をこえない)
-
さらに行動連鎖のなかで先に antennation をやってしまうと,
(観察者には) 攻撃確率が下っているように見える
……
といったところが中軸だろうか,
と.
つまり何かにシロクロつけるハナシではなく,
アリの行動もっとよく見て
(行動連鎖とか),
きちんと統計モデリングしなさい,
というハナシになりそう.
-
とアタマの中で検討しつつ,
``Polydomy in ants: ...'' (2007) 総説論文よみのつづき.
時間かかるけど,
こういう (ふだんでいりしてない) 分野の総説論文よみは
ゆっくりメモとりながら読みススめていくほかない.
-
``Aggression test conducted using pairwise reciprocal
comparisons of mature nests, ...''
ですか.
ちっとも reciprocal じゃない敵対性実験データの解析しているんだけどなあ,
と思いつつひと区切りつけて昼飯.
-
夕方までかかって,
どーにかこーにかこの総説ひととーり読めた.
多巣性 (polydomy) ってのがどういうものか
(そしてそれを記述する文献は何か),
といったことはよくわかるけど,
polydomy のご利益などといった生態学的な研究は何もススんでおりません,
というかんぢだ.
こういう「個体数の変化」にまつわるハナシはアリ研究の弱点で,
アリ科研費したうけぎょーむにおいても
慎重に回避すべきところなんだろう
……
ということで,アリシミュレイション構築は放棄した.
Polydomous なアリの動態は複雑なんで,
抽象化も難しくしたがって進化なハナシも困難だろう.
-
ちなみにこの総説の Table 1 でエゾアカヤマアリ
(Formica yessensis)
はこうなってる
(出典はほとんど Keller ed. (1993) Queen number spciality in insects 本):
-
Distribution = PAL (Palearctic) (Japan, Korea)
-
Climatic region = TE (Temperate)
-
Habitat = TF (Temperate and boreal forest)
-
Nest site = T (Terricolous)
-
Nest type = C (Self-construction)
-
Ecological status = D (Dominant)
-
Type of polydomy = F (Facultative (size dependent))
-
Seasonal polydomy = n (No)
-
Associated gyny = P (Strict polygyny)
-
Colony size = 8 (> 106)
-
エゾアカヤマアリ原稿のみなおし,
ちょっとだけ.
bibtex で修士論文 (修論) を引用するときの entry
のかきかた.
@Masterthesis
なる entry 名もあるんだけど,
ちゃんと動作してるように見えないので,
@Phdthesis{Iwakura2007,
Author = {Misako Iwakura},
Title = {
Effects of genetic differentiation and cuticular hydrocarbonic composition
on internidal aggressiveness in a supercolonial ant
\textit{Formica yessensis}
},
School = {Graduate School of Environmental Science, Hokkaido University},
Type = {{Master} thesis},
Year = 2007
}
というふうに @Phdthesis entry + Type
わざ,
でそれっぽく表示される.
-
えーい,
先方の都合で順延したアリモデリング談判の再開は当方からもうしこめ,
というのが大統領府の方針なのかしらん?
まあ,
しばらく放置.
-
1930 研究室発.
1945 帰宅.
体重 68.0 kg.
晩飯の準備.
晩飯.
-
[今日の運動]
-
腹筋運動 30 ×
3 回.
腕立ふせ 10 ×
3 回.
スクワット 100 回.
-
ラジオ体操.
ストレッチ.
-
[今日の食卓]
- 朝 (0830):
米麦 0.5 合.
ネギ卵炒飯.
キャベツ・ニンジン・タマネギ・キュウリ・レタスのサラダ.
- 昼 (1310):
研究室お茶部屋.
食パン.
キャベツ・ニンジン・タマネギ・キュウリ・レタスのサラダ.
- 晩 (2210):
米麦 0.8 合.
ダイコンの味噌汁.
キャベツ・ニンジン・タマネギ・キュウリ・レタスのサラダ.
ピーマン・ネギ・ダイコンの皮・ブナピー・ショウガ・
ニンニク・卵の炒めもの.
2007 年 07 月 10 日 (火)
-
0805 起床.
コーヒー.
朝飯.
0905 自宅発.
晴.
0920 研究室着.
-
セミナーの予習.
-
1030 より
研究室セミナー,
本日は稲葉さんで,
植物が (昆虫とかに) くらった損傷と abortion (種子形成放棄)
の関係とか.
下記 4 論文のうち,
上から 3 論文についての検討.
- Spotswood et al. (2002) Effects of herbivory on the reproductive effort
of 4 prairie perennials. BMC Ecology 2 (2)
Background: Herbivory can affect every aspect of a plant's life.
Damaged individuals may show
decreased survivorship and reproductive output.
Additionally, specific plant species (legumes) and
tissues (flowers) are often selectively targeted by herbivores, like deer.
These types of herbivory influence a plant's growth and abundance.
The objective of this study was to identify the effects of
leaf and meristem removal (simulated herbivory within an exclosure) on fruit and flower
production in four species (Rhus glabra, Rosa arkansana, Lathyrus venosus, and
Phlox pilosa) which are known targets of deer herbivory.
Results: Lathyrus never flowered or went to seed, so we were unable to detect any
treatment effects. Leaf removal did not affect flower number in the other three
species. However, Phlox, Rosa, and Rhus all showed significant negative correlations
between seed mass and leaf removal. Meristem removal had a more negative effect than
leaf removal on flower number in Plhilox and on both flower number and seed mass in Rosa.
Conclusions: Meristem removal caused a greater response than defoliation alone in both
Phlox and Rosa, which suggests that meristem loss has greater effect on reproduction.
The comibation of leaf and meristem removal as well as recruitment limitaion by deer,
which selevtively browse for these species, is likely to be one factor contributing
to their abundance in prairies.
- Marcelis et al. (2004) Flower and fruit abortion in sweet pepper in
relation to source and sink strength. Journal of Experimental Botany
55 (406) 2261-2268
(abstract)
Source strength (assimilate supply) and sink strength
(assimilate demand) of the plant were varied in different
ways to investigate to what extent flower/fruit abortion
in sweet pepper (Capsicum annuum L.) is determined by
the availability of assimilates. Source strength was
varied by changing the light level, plant density, and
leaf pruning. Sink strength was varied by changing the
temperature and the number and position of earlier
formed fruits. Shading as well as heating for short
periods showed that when source strength decreased,
the rate of abortion increased linearly, whether source
strength was decreased by shading, high plant density,
or leaf pruning. That flower and fuit abortion not only
depends on the source strength but also on the sink
strength of competing organs is shown by varying the
number or the position of earlier formed fruits. With the
same source strength, the rate of abortion showed
a close relationship with the growth rate of the earlier
formed competing fruits, suggesting that the induction
of abortion by earlier formed fruits is due to their sink
strength. Most of the variation in abortion could be
related to differences in vegetative growth rate, the latter
being an indicator or the source-sink ratio. However,
with the same vegetative growth rate, the rate of abortion
was lower for the leaf pruning treatments where no
competing fruits were retained than for the fruit load
treatments. This indicates that although most of the
variation in abortion can be related to the source and
sink strength of the plant, some effects of competing
fruits can only be explained by a combination of competition
and dominance.
- Hellstrom et al. (2006) Plasticity of clonal integration in the
perennial herb Linaria vulgaris after damage. Functional Ecology
20 (3) 413-420
1. Clonal integration in plants can improve their ability to cope with heterogeneity.
Integration may increase in response to damage, such as herbivore attack, if
undamaged ramets support damaged ones. To test this, we studied the effect of apex
removal and substantial defoliation on the perfomance of the clonal perennial herb
Linaria vulgaris Mill. in a common-garden growth experiment and 13C-labeling
study.
2. In the growth experiment, contrary to expectations, the target ramet could compensate
for damage better wht the other ramets in the clone were also damaged, indicating
within-clone competition for resource rather than support to damaged ramets.
3. In the 13C-labeling experiment, 5-7% of the lable moved to a neighbor ramet
in controls. Apex removal resulted in a negative net translocation of 13C in
the damged ramet, but defoliation led to zero net translocation.
4. The observed lack of support to damaged ramets in Linaria suggests that plasticity
of clonal integration in this species includes competition between sibling ramets.
Although young ramets may be supported, resources are not directed towards a single
damaged ramet if there are more viable intact ramets in the clone. Our main results
are consistent with the notion that resource allocation among ramets depends on their
relative value in term so fexpected fitness profits in heterogeneous environments.
- Ishikawa et al. (1990) Photosynthetic characteristics and water use
efficiency of three coastal dune plants. Ecological Research 5 (3) 377-391
To clarify the ecophsiological characteristics of plants growing on a coastal dune,
net photosynthetic rate (Pn) and leaf conductance (g1) of three perennial species in
Japanese coastal regions, ...
were compared under controlled environmental conditions and field
conditions at the Kado-ori coast at Ohno Village, Ibaraki. I.anthepheroides achived
photosynthetic CO2 saturation at ca. 100 μl-1 intercellular CO2 concentration (C1),
andits Pn was not light-saturated at a high photosynthetically active photon flux density
(PPFD) of 1000 μmol m-2 s-1. This C4 species showed a high optimal leaf temperature
for Pn (35°C) and a low g1 (0.1 mol H20 m-2 s-1), permitting maintenance of the
highest water use efficiency (WUE, the ratio of Pn to transpirationrate (Tr)) in the field
among the three species.
...
...
ハナシは多年性草本のシュート・ramet における
「今シーズンの繁殖からの撤退を決定づける損傷」
は何か,
といった方向のもので,
葉がやられるのはとうぜん痛いんだけど,
apex だの meristem がヤラれると
(たぶんシグナル分子とかの関連で)
sink 機能低下するようで
「今年はやめた」とあきらめてしまうらしい,
と.
あきらめると隣のシュートに同化物をぶんどられたり
(上から 3 番目の論文),
あるいはもしかしたら地下部にまわして来年用にしてるかも,
と.
-
う,
のんびりと議論やってたらセミナー終了が 1250 になってしまった.
昼飯.
-
お茶部屋で亀山さんと
「ある雑種らしき植物個体の遺伝子 (species specific な遺伝子座)
みたときに,その個体が F1, F2,backcross のいずれかを判定する」
統計モデリンングについて検討.
ようするに
-
F1 だの何だのという個体ごとの labeling
(もしくは assignment)
-
species specific ということになってる遺伝子の頻度
(これは一個体だけでなく標本集団全体で決まる)
のどちらも不確定で,
片方を決めるともう一方が変わる
……
ということで Bayesian とかであつかうべき典型的な問題である.
しかしここでまた新しい階層ベイズモデルなど発明してしまうと
論文一本ぶんの仕事になってしまい
……
(とある事情によってこの問題を安直に解決しなければならぬ現状では)
ちょっと大げさすぎるのである.
-
ということで,
可能なかぎり手ぬきする方法を考えてみた
……
うーむ.
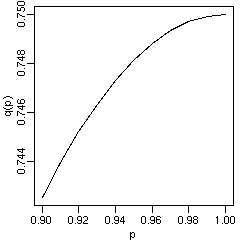
[F2 は遺伝子頻度の影響が少ない]
しかしいっしょに検討してると,
いろいろなことがわかった.
ある「種特異的」対立遺伝子がわりと高めの頻度 p で存在するときに,
F1 におけるその遺伝子のあり・なしは p の影響をもろにうけるけど,
F2 になると dominance の効果で応答性が鈍感になる,
とわかった.
-
1600 から農学部の徐さんのデータ解析こんさる.
-
1700 ごろからまた亀山さんと雑種問題の検討.
実データをつかって
「F1 であるという尤度」
「F2 であるという尤度」
計算させてみたんだが (くだんの遺伝子頻度をてきとーに与えて),
まあいくつか問題のある個体が.
しかもそれが遺伝子頻度に影響されてて,
すっきりしない.
-
とうとうここでも
「めんどうになったらベイズ」
という近ごろのおきまりのパターンが適用されて
……
驚くべきことに
「ある個体が F1 なのか F2 なのかわからん,
ホントの遺伝子頻度もわからん,
それでもともかく個体 i が F2 である確率を求めよ」
という限定的な問題を解決する統計モデルは
ゐんばぐす
で下のごとくごく簡単に記述できる,
とわかった
……
超事前分布すら不要!
# N.sample 個体の遺伝子データ,遺伝子座数は N.loci
# 個体 i の遺伝子座 X[i, j] は 0, 1 のデータをとる (AFLP データ)
# 個体 i が F1 であるとき isF2[i] <- 0,
# F2 であるとき isF2 <- 1 (unknown)
# 遺伝子座 j における種特異的対立遺伝子の存在確率は p[j] (unknown)
model
{
# F1 vs. F2 アサインメント
for (i in 1:N.samples) {
isF2[i] ~ dbern(0.5) # 0, 1: 無情報事前分布
type[i] <- isF2[i] + 1 # 1, 2
for (j in 1:N.loci) {
X[i, j] ~ dbern(prob[type[i], j])
}
}
# 種特異的遺伝子の頻度
for (j in 1:N.loci) {
p[j] ~ dunif(0, 1) # 無情報事前分布
prob[1, j] <- p[j]
prob[2, j] <- 1 - (
p[j] * p[j] / 4 + p[j] * (1 - p[j]) / 2
+ (1 - p[j]) * (1 - p[j])
)
}
}
いつものごとく,
これを
R2WinBUGS
で計算させてみると,
500 MCMC step ぐらいで十分に収束.
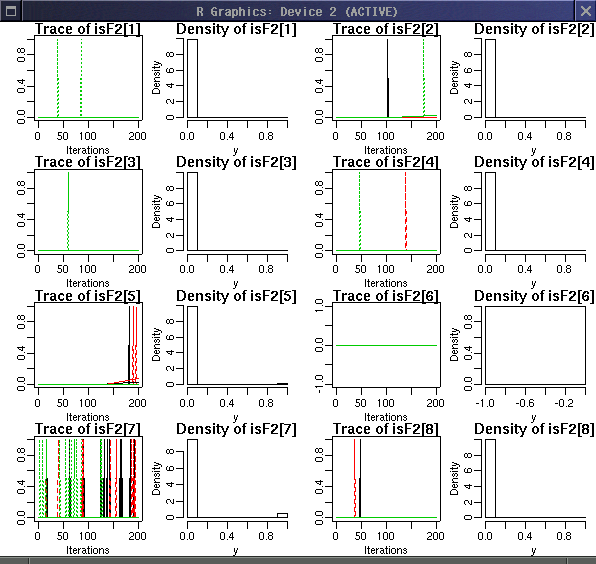
個体 i が F2 かどうかをあらわす isF2[i]
の事後分布.
0 と 1 のあいだをいったりきたりしてる.
-
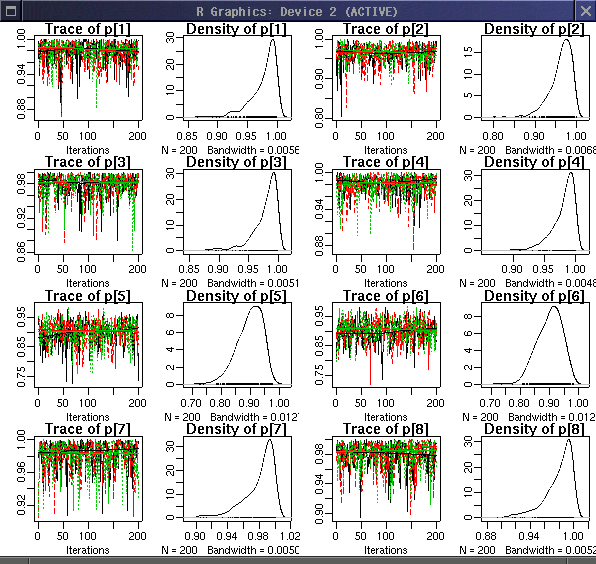
そして unknwon な遺伝子頻度
p[j]
の事後分布.
-
……
で問題はいちおうの決着をみた.
しかし作ってみると,
「ハイブリッド問題 (というかアサインメント問題)
に関して Bayesian がこれほど強力な道具であるならば,
だれかがこういう計算プログラムをすで作って公表しているにちがいいない」
という集団遺伝学まわりの分野の良い文化にもとづく推測が成立するので
……
検索してみると,
やはり見つかった.
Eric Christopher Anderson の
NewHybrids,
Anderson, E.C. and Thompson, E.A. (2002) A model-based method for identifying species hybrids using multilocus genetic data. Genetics 160: 1217--1229
(上記 URL から PDF file ダウンロード可能).
-
ということで,
亀山さんはこの NewHybrids でもって F1 vs. F2 だけでなく,
バッククロスも含めたより包括的な問題にとりくむことに.
2110 研究室発.
晩飯.
2220 帰宅.
-
[今日の運動]
-
[今日の食卓]
- 朝 (0820):
米麦 0.5 合.
ダイコンの味噌汁.
キャベツ・ニンジン・タマネギ・キュウリ・レタスのサラダ.
ピーマン・ネギ・ダイコンの皮・ブナピー・ショウガ・
ニンニク・卵の炒めもの.
- 昼 (1310):
研究室お茶部屋.
食パン.
キャベツ・ニンジン・タマネギ・キュウリ・レタスのサラダ.
- 晩 (2140):
北 13 西 3 のピカンティ (スープカレー専門店),
大陸 X スープのベイコン with キャベツ.
集団遺伝学ベイズモデルの
プログラミングばてのカラダにはいいかんぢでした.