ぎょーむ日誌 2007-05-(21-31)
2007 年 05 月 21 日 (月)
-
0700 起床.
コーヒー.
朝飯.
0820 自宅発.
晴.
0835 研究室着.
-
地衣類繁殖の志水さんデータ解析の階層ベイズ説明
……
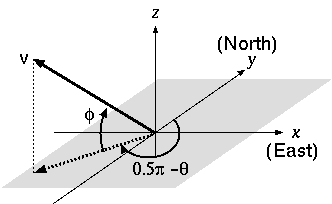
の手前の太陽軌道説明図かきあれこれ.
-
三次元作文.
正午前にばててきたので昼飯.
-
三次元作文のつづき.
匍匐前進的.
-
データ解析こんさる.
こんにちの群集生態学 (ようするに多種系の
…… まあ,構造をあつかうデータにたちむかうもの)
でつかわれてるいわゆる「多変量解析」
ってつくづく変てこな世界だよなぁ
……
まあ,
今後はどんどん変わっていくだろう.
もちろんマシな方向にむかって,
だ.
-
今日のところは従来方式をこえられなかった挫折感つのってきたので,
北大構内散歩.
たいへんさわやかな天気.
-
北大構内あるいてるうちに,
太陽軌道三次元計算の一番めんどうな部分を思い出すことができた
……
「思い出す」ってのは,
昨年 10 月
の時点で考えついたことはちゃんとメモしといたんだけど,
一式ぜんぶ志水さんに渡してしまっていて
(そして志水さんは「本業」(?) 多忙で不在の日々),
手もとになかったんだよね
(ただし上のような図を作図できる太陽軌道計算 R コードだけが残っていて,
ですね).
回転は下のごとし.
[x 軸を回転軸とする回転]
ようするに
「x 軸を『回転軸』として y-z 平面をまわすんだけど,
各点はとうぜんながら『球面上』を移動していく」
というだけのことだ.
つまり各点ごとに「どの」 y-z 平面に乗ってるのか注意しなさい,
ということ.
なんで最初に
青
みたいなヘンな架空の太陽軌道を準備するかって?
これを出発点にしたほうが便利だからだよ
……
いや,
やっぱり,
わかりにくいかな
……
-
ということで,
赤道からみた黄道を基準にして,
これを「y-z 平面で回転」させることにする.
-
1715,
どーにかこーにかひととーり三次元計算説明かけた
……
-
誰がこんな三次元くるくる回転計算なんかいちいちチェックするかっつーの,
てなかんぢでしょうか.
まあ順当に Appendix 送りなんでしょうな.
そして
oowrite
(おうぷんおひす
のわーど)
の数式・レイアウトは見苦しい
(そして数式が増えるにつれ処理が激オモに).
LaTeX
でサクサク美しく書きたいようー
-
とりあえず白背景は目がおかしくなりそうなので
(皆さんよくガマンできますよね),
グレイにする.
いやー,
いつも
こういう暗色
(google image 検索)
の画面みてるもんで
……
-
三次元計算説明から脱出して階層ベイズモデル説明かき.
こちらの進捗も匍匐前進的だけど,
アタマの中でヴェクトルぐるぐるまわさなくてすむのでラクだ
……
なるほど,
(地衣類の)
子器の単数形が
apothecium
で複数形が apothecia,
なのか.
-
おうぷんおふぃすもやや不安定だなあ.
印刷出力命じたらふりーづしたよ.
しかも強制終了しても background にはたくさんの
soffice.bin
走ってるし
……
こんなの (ゐんどーづ世界とかで暮らしてきた)
一般ユーザーは対処できんよ.
-
また事前分布を列挙するのが面倒になったので,
いつものごとく Table に書いて始末することに.
しかしおうぷんおふぃすで Table なんか作ったら
胃のあたりがキモチ悪くなりそうだから,
LaTeX で作ることにしよう.
Figure はもとより
R
/
Tgif
+ LaTeX で作りつつある.
問題は References だなぁ
……
まあモデル部分は引用文献がほとんどないから気にしなくてよいか.
-
Table は LaTeX で
……
嗚呼,
キモチいい
……
-
ばててきたので撤退.
2020 研究室発.
2035 帰宅.
晩飯の準備.
晩飯.
-
[今日の運動]
-
腹筋運動 30 ×
3 回.
腕立ふせ 10 ×
3 回.
スクワット 100 回.
-
[今日の食卓]
- 朝 (0730):
米麦 0.6 合.
鶏レバの煮物.
- 昼 (1150):
研究室お茶部屋.
米麦 0.6 合.
春雨担々麺カップスープ.
- 晩 (2140):
米麦 0.8 合.
ニラ・ピーマン・卵の炒めもの.
タマネギ・ジャガイモ・ニンジンの味噌汁.
2007 年 05 月 22 日 (火)
-
0710 起床.
コーヒー.
朝飯.
0810 自宅発.
晴.
0825 研究室着.
-
セミナー予習.
-
1030 より
研究室セミナー,
本日は矢澤さんで熱帯林論文紹介.
-
Falster, D. S., and M. Westoby. 2005. Alternative height strategies
among 45 dicot rain forest species from tropical Queensland, Australia.
Journal of Ecology 93:521-535.
1. Potential height, which spans at least an order of magnitude across species,
is considered an important indicator of light capture strategy. Still it remaines
unclear how potential height is coordinated with other traits that influence height
growth rate, stem persistence and performance in low light. We proposed that contrasting
correlation between potential height and other plant attributes would be observed for
sets of species selected to span two hypothetical axes of light availability within
mature forest and time since disturbance.
2. We selected 45 perenial rain forest species in Australita's wet tropice to span
gradients of light availability and successional status and measured potential height
together with traits inlucencing light capture and regeneratrion strategy on mature
individuals. The traits included leaf mass per area, leaf nitrogen, wood density, stem
mass per length, branch mass fraction and seed mass.
3. Potential height was significantly correlated with numerous traits among species
selected to span each of the two gradients. Height was positively correlated with
leaf mass area, leaf nitrogen and seed mass and negatively correlated with leaf area
ratio at the branch tip along both light and successional gradients. Height was
positively correlated with wood density along the successional axis, with the opposite
relationship along the light gradient.
4.The relationships differed in either slope or intercept between two gradients,
reflecting different strategic trade-offs. At a given height, shorter species in the
succesional gradient were chracterized by lower leaf mass area, lighter wood, smaller
seeds, lower leaf nitrogen and lower leaf area ratio at the branch tip than similar
sized species along the light gradient.
The results of this study support the idea of two distinct, trait-mediated axes of
coexistence among short and tall plant species within vegetation. In several cases,
trait relationships were weak or non-significant when species groupings were merged,
indicating the importance of separating out the two sets for comparison studies.
-
Kohyama, T., E. Suzuki, T. Partomihardjo, T. Yamada, and T. Kubo. 2003.
Tree species differentiation in growth, recruitment and allometry in
relation to maximum height in a Bornean mixed dipterocarp forest.
Journal of Ecology 91:797-806.
-
Poorter, L., F. Bongers, F. J. Sterck, and H. Woll. 2003. Architecture
of 53 rain forest tree species differing in adult stature and shade
tolerance. Ecology 84:602-608.
The architecture determines a tree's light capture stability, and efficiency
of crown growth. The hypothesis that light demand and adult stature fo tree species
within a community, independently of each other, determine species's architectural traits
was tested by comparing 53 Liberian rain forest tres species. We evaluated whether species
differed in there tree height, crown depth, and crown diameter, whne compared at a
standarized size of 15-cm diameter at breast height, and how their architecture changed
early during ontogeny.
Tree height was positively correlated with adult stature and light demand. By producing
a relattively slender stem, large stature species area able to rapidly reach their
reproductive size, at a low cost for construction and support. Light-demanding species
need a slender diameter were negatively correlated with adult stature, but not with
light demand. This is in cotrast with the hypothesis that shade-tolerant species should
have a shallow crown to reduce self-shading in a light-limited environment. Investigating
energy in height growth rather than lateral crown growth allows a rapid vertical stem
extension, but crown diameter has to be sufficiently small to reduce the risk of
mechanical failure. All architectural patterns were maintained during ontogeny. The key
factors driving interspecific differences in tree architecture are the costs of height
extension and mechanical stability. In general, light demand and adult stature represent
independent axes of architectural differentiation, affecting tree architecture in
different ways.
-
Sterck, F. J., and F. Bongers. 2001. Crown development in tropical rain
forest trees: patterns with tree height and light availability. Journal
of Ecology 89:1-13.
1. Monitoring of two canopy species Dicorynia guianensis and Vouacapoua americana
(Casesalpiniaceae) in tropica rain forest in French Guiana was used to investigate
vegetative crown development at five organizational levels: leaf, metamer, extension
unit, sympoidal unit and whole crown. The effects of light availability and tree height on
different traits were evaluated in trees < 25 m in height and compared with taller
individuals (25-37 m). Path analysis is used to illustrate the consequences of traits
changes at multiple levels of organization for the whole crown level.
2. Tree height and canopy openness influenced crown development at each organizational
level. Crowns in higher light levels had lower specific leaf area, greater leaf spacing,
greater extension of all branches, and greater extension of the leader shoot. With
increasing tree height, crowns had a lower specific leaf area, greater leaf area index
and greater relative crowth depth.
3. Vouacapoua showed some responses to light not seen in Dicorynia. In particular,
Vouacapoua increased meristem activity with light, but the lack of reponse in Dicorynia
may be due to moderate light levels rather than iablity to repond.
4. Low leaf-diplay costs at low light availability may enable trees to survive light
suppressions.
5. Light availability cannot explain trait changes with tree height. Alternative
explanations for trait changes with tre height are discussed.
6. Several of the relationships between plant traits adn tree height or canopy openness
became non-linear when taller trees (25-37 m) were included. In these taller trees,
vegetative growth was reduced at all organizational levels, particularly in Vouacapoua,
which does not grow as Dicorynia.
7. Qualitatively, plant responses to light did not differ between trees of different
height, and were similar to seedling and sapling data in the literature. Responses were,
however, qunatitatively different, suggesting that small saplings cannot serve as model
organisms for crown development in taller trees.
-
Thomas, S. C. 1996. Asymptotic height as a predictor of growth and
allometric characteristics in Malaysian rain forest trees (vol 83, pg
556, 1996). American Journal of Botany 83:1570-1570.
うーむ,
どれもわざわざ熱帯まで行ってやる必要のない研究ばかり,
という気もするんだけど.
-
1220 終了.
1230 昼飯調達のため北大構内の旅にでる.
今日もいい天気.
中央ローンは戸外で昼飯たべてるヒトたち多数.
北大生協うろうろしてから研究室もどって昼飯
……
食おうとしたんだけど,
お茶部屋の院生密度が高くてなかなか近づけない.
-
などとうだうだしてるうちに,
7F 実験室の雑用 PC につけてる CRT ディスプレイ壊れ連絡が
……
うーむ,
私が D3 のときに買った X 端末専用機
(というのが当時あった)
についてたやつじゃないか.
そりゃ壊れますわな.
新たなる粗大ゴミ一個をまた一時ゴミ置場 (つまり私のいる部屋)
に.
JCC 製.
驚くべきことに,
この
したたかな
計算機会社は過去 10 年間の激動の時代を生きのびてしまって,
いまだに存続している,
とわかった.
何だか感慨ありますねえ
……
-
ということで 1400 ごろ昼飯.
-
昼飯後にしばらくうだうだとしてから,
また地衣類子器数
の階層ベイズモデルの説明かきつづき.
-
しかしつくづく「わーぷろ」
(おうぷんおひすのらいたー)
にはいらいらさせられる,
というほどでもないけど楽しくない
……
LaTeX はあんなに楽しいのに.
-
1700 すぎ,
ひととーりモデル説明とか書けたかしらん,
などと思ってると
……
後藤さんから改定原稿一式がおくられてきてトドマツ世界にひきもどされる.
-
1850 研究室発.
1905 帰宅.
晩飯.
-
2210 自宅発北大構内走.
ゆっくり走って
2300 帰宅.
-
[今日の運動]
-
[今日の食卓]
- 朝 (0730):
米麦 0.6 合.
タマネギ・ジャガイモ・ニンジンの味噌汁.
- 昼 (1400):
研究室お茶部屋.
食パン.
チーズ.
- 晩 (1920):
米麦 0.8 合.
寄せ豆腐.
タマネギ・ジャガイモ・ニンジンの味噌汁.
2007 年 05 月 23 日 (水)
-
0750 起床.
コーヒー.
朝飯.
0930 自宅発.
晴.
0940 研究室着.
-
さいきんの
Ecology Letters
は毎号のように Bayesian & MCMC な論文が掲載されてるな
……
これもおもしろそうだ.
Subhash R. Lele, Brian Dennis, Frithjof Lutscher.
2007.
Data cloning: easy maximum likelihood estimation
for complex ecological models using Bayesian
Markov chain Monte Carlo methods.
Ecology Letters (OnlineEarly Articles) 10:???-???.
doi:10.1111/j.1461-0248.2007.01047.x
-
数ヶ月前に,
とある (やや工学より) proceedings の査読やったら Amazon
ギフト券 3000 円もらってしまった.
さすがは「やや工学より」,
カネもちだねえ.
ということで,
さっそく Bayesian data analysis 本購入にぽーんとつかってしまった.
-
トドマツ原稿にとりくむ.
そうだな,
まあ discussion 節の改定文案かきから始めるか.
-
えーと,
当方愛用のテキストエディター
Vim で dos な (Shift-JIS)
なファイルきめうちで開くには
……
vim "+e ++enc=cp932 ファイル名",
と.
-
いつものごとく遅々たる作文を 1230 ごろまで.
さーて,
作文はいったんおいて,
また事後分布まわりの計算をやらんといかんね.
まあ,
これがひかえめにいっても面倒きわまりないもので.
-
いきなり 30 分ほど 7F に拉致されて,
甲山さん・山中さんの Global COE ヒアリング練習につきあわされることに.
明日が本番だそうで.
-
ちょっと計算すすめてから昼飯.
-
また散布距離分布問題.
R
で (事後分布から MCMC sampling された標本の)
``quantile'' 値だす方法いろいろありそうだけど,
おそらく一番簡単には,
quantile()
使う方法で,
たとえば 95% 区間はこう計算される.
> x <- rnorm(100000, 0, 1) # ここでは例題としててきとーな乱数
> quantile(x, probs = c(0.025, 0.975))
2.5% 97.5%
-1.9662 1.9530
-
さて,
「平均散布距離」とやらの 95% 区間だけど
……
79 母樹それぞれに 600 個の (事後分布からの) 標本値があるので,
「事後」散布カーネルは合計 47400 個あるわけだ.
とうぜん,
それぞれが平均値 (
sqrt(0.5 * pi) * sigma)
もつわけで,
その中央値 (50% 値) と 95% 区間はこうなる.
normal2D(sd0) 33.6 (28.2, 41.3)
normal2D(sd0 + sdMt) 31.3 (7.2, 210.6)
上の
normal2D(sd0)
が (LMM ではなく GLMM 的な意味あいでの) ``random effects''
ぬきモデルで (sdMT <- 0;
蛇足ながらこいつの平均値は 34.0 m),
normal2D(sd0 + sdMt)
は 79 母樹差 random effects ありモデル
(蛇足ながらこいつの平均値は 51.6 m).
後者は,
いやはや,
ナイスな混合モデル的ばらつきかたですね.
-
とびこみ
R
こんさる.
「Y 軸のラベルを水平にしたい」
ですか.
しかもよりによって「ぼうぐらふ」とは
…… !
ともあれ回答例送信.
x <- 1:10
names(x) <- LETTERS[x]
par(las = 1)
barplot(x, horiz = TRUE)
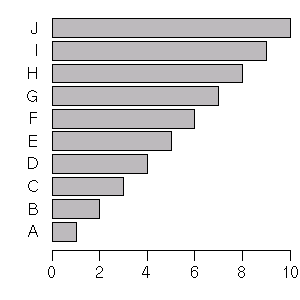
[とりあえず help(par()) を]
par() の help
はなかなかごちゃごちゃしてるんだけど,
horizontal とかで検索すればすぐに
par(las = ...)
にいきあたります,
と.
-
トドマツ原稿 discussion 節かきなおしつづく.
-
現実逃避というわけでもないけど,
(院生密度がなぜかひくいので)
お茶部屋で亀山さんややのんびりとツガザクラ雑談.
いろいろ発見が続いてますねえ
……
ところで,
雑種 F1 はできるけど F2 ができない,
という現象を説明するには
複数遺伝子座の相互作用を考えないといけないのだろうか?
-
またトドマツの
paper/section/discussion.tex
の修正作業などなどを続けて
……
まあ,
本日はこんなところかと
make pdf && make save
の rsync
でバックアップして,
「こう変えてみました」メイルかいて送信.
時刻はすでに 1855.
-
1940 研究室発.
1955 帰宅.
体重 67.6kg.
-
晩飯後に原稿にとりくんでみたんだが
……
仕事すすまん
……
-
[今日の運動]
-
[今日の食卓]
- 朝 (0840):
米麦 0.6 合.
納豆.
ナメコの味噌汁.
- 昼 (1400):
研究室お茶部屋.
食パン.
チーズ.
リンゴ.
- 晩 (2030):
米麦 0.8 合.
ニラ・エノキダケ・卵の炒めもの.
2007 年 05 月 24 日 (木)
-
0740 起床.
コーヒー.
朝飯.
0915 自宅発.
晴.
0930 研究室着.
-
トドマツ原稿ファイル修正作業.
-
昼前に東大日光植物園の市橋さんがきてくださったんだが
……
誰も対応するヒトがいないので,
宮田さんにおともをお願いして,
私が昼飯におさそいすることに.
ツル植物の楽しいハナシをいろいろとうかがう.
-
1330 より
Trendy セミナー,
市橋さんによる木本 (落葉) ツル植物の生態学 talk.
サルナシ,ツルウメモドキ,マツブサ,ミツバアケビ,イワガラミ
(←これだけ付着根もつもので,他の 4 種は巻きつき茎型)
の 5 樹種の比較.
興味ぶかかった点としては
……
-
ツル植物にもシュートの機能分化あり,
探索枝・普通枝にわかれる
……
これはほぼ長枝・短枝に対応?
(普通枝から探索枝はでない,
など)
-
さらに探索枝重視か普通枝重視かで
森林内のすみ場所がちがっていそう
……
探索枝重視のサルナシなどは林縁の (樹高が比較的低い)
host 木にとりつき
(そして樹冠上に葉を展開),
普通枝重視のものは林内の host 木に
(そして樹冠内に葉を展開)
-
さらにサルナシみたいなツル植物は「移動」して
host 木を乗り移っていくらしい
……
目的論的な説明としては林縁ではよく倒木するから

[ツル植物セミナー]
この生活形の植物を
モデル化して (つまり抽象的に) 考えると,
ふつーの木本樹木のように考えることもできそうな部分
(長枝・短枝あるいは伸長・繁殖への資源分配,
光・水ストレスに応じた伸長様式など)
とつる植物独特な部分 (探索枝をたくさんだすと
host 木を変更できる可能性たかまるので寿命を延伸できる)
をわけたらおもしろいかも,
と.
-
低温研の黒沢さんもセミナーに参加されていて,
ツルウメモドキまにあっくなハナシを
(カラスは種子散布者かもしれないけどフンが大きすぎると
発芽直後の高密度状態にツルウメモドキが耐えられず自滅するかも?
など).
昨年 10 月
の John Silander さんのハナシともいろいろと関連していたり.
あと,
今回の 5 樹種の中で,
唯一ツルウメモドキだけは (シウリザクラのごとく)
根萌芽 (sprouting) するそうだ.
-
その後,
お茶部屋でまたツル植物雑談.
永光さんもセミナーにきておられたので,
シウリザクラ相談なども.
今日はひさしぶりに口頭による生態学ハナシをいろいろやった,
というかんぢだ.
-
またトドマツ原稿作業.
-
本日なすべき修正作業はひととーり終えて,
また一連のファイルアップロード作業.
1845 研究室発.
1900 帰宅.
洗濯.
晩飯の準備.
晩飯.
-
2300 自宅発北大構内走.
2345 帰宅.
-
[今日の運動]
-
[今日の食卓]
- 朝 (0800):
パンケイキ.
- 昼 (1230):
市橋さん・宮田さんと「きゃら亭」の昼定食.
白身魚 (なんだったのだろう?) ソテー,
和風ソース.
- 晩 (1930):
米麦 1.0 合.
ニラ・豆腐のトマトスープ.
2007 年 05 月 25 日 (金)
-
0830 起床.
コーヒー.
朝飯.
0930 自宅発.
曇.
ちょっと寒い.
0945 研究室着.
-
うだうだとしてから,
またぢりぢりとトドマツ原稿にとりくむ.
今日は (discussion の内容も限定されてきたので)
モデル説明まわりの作文改善,
とか.
しかし作文しようとすると,
いろいろトドマツ質問メイルをいただいたり.
-
うーむ,
散布カーネルな確率分布の平均値って,
``tail'' 部分に敏感だろう,
と.
そして調査 plot を広げれば広げるほどますます
「遠方に散布されてしまった子供」が見つかったりするので,
それにつれて
「平均散布距離」はいよいよ延伸されてしまうかも,
と.
-
そして「random effects とは何か?」
の定義に関して「一般化した」説明をまたひとつ思いついた.
平均値にもとづく説明はヤメて,
「分布の中央値は変えないけれど,
ばらつきを変えるのが random effects」
というのはどうだろう?
これに対して fixed effects は中央値を変える,
と.
おそらくこれで consistent なハズなんだけど,
わかりやすくないので
(つまりふつーは確率分布を中央値で考えないので)
あまり多くの人から支持されないような気もする.
-
作文なおしは着手できぬまま昼飯.
-
R-News
の最新号 (Volume 7/1, April 2007)
に ``A New Package for Fitting Random Effect Models''
として
library(npmlreg)
(Nonparametric maximum likelihood estimation for random effect models;
library(help = npmlreg))
が紹介されてるな
(引用太字は久保による):
Random effects
have become a standard concept in statistical modelling over
the last decades. They enter a wide range of applications by
providing a simple tool to account for such problems as model
misspecification, unobserved (latent) variables,
unobserved heterogeneity, and the like. One of the most important
model classes for the use of random effects is
the generalized linear model.
Aitkin (1999) noted that
``the literature on random effects in generalized linear models
is now extensive,'' and this is certainly even more true today.
で,
ここでいう nonparametric maximum likelihood は random effect parameters
の分布を決めずに (たとえば
glmmML()
だとこれを正規分布と仮定している),
The random effects distribution can be considered as an
unknown mixing distribution and the NPML estimate of this
is a finite discrete distribution. This can be determined
by fitting finite mixture distributions with varying numbers
of support points, where each model is conveniently fitted
using a straightforward
EM algorithm.
……
ということで,
EM algorithm
つかった GLMM の推定 package なのである.
ふーむ,
またこんど使ってみるか
……
-
8 月のアリ研究会,
事務とりまとめの三浦さんからのキビしい追及
(その背後に
executive order?)
から逃れられなくなり,
「アリデータの統計モデリング」
なるいいかげんな講演タイトルをだしてしまう
(しめきりは 5/31 だったはずなのに……).
さっさとトドマツと地衣類繁殖の作文ぎょーむをかたづけてしまわなくては
……
-
と思ったら原稿チェックのすばやい練さんから
「ここが変ですよ」
ご指摘一覧が.
うう,
私が昨日追加した作文も含まれている
……
-
修正してアップロード.
時刻はすでに 1700 ちかい.
-
トドマツ原稿修正のほうはあまり進捗せず
……
1800 研究室発.
雨.
1825 帰宅.
晩飯の準備.
晩飯.
-
Wired Vision
のりにゅーあるが進捗している
(Hotwired からのお知らせ).
-
[今日の運動]
-
[今日の食卓]
- 朝 (0850):
パンケイキ.
ニラ・豆腐のトマトスープ.
- 昼 (1330):
研究室お茶部屋.
食パン.
チーズ.
リンゴ.
- 晩 (2020):
米麦 0.7 合.
ネギ・ジャガイモ・ニンジン・ブロッコリー・ブナピー・
干しエビ・ホタテのクリームシチュー.
豚タン・タマネギ・プチトマト・レタスのサラダ.
キュウリ.
2007 年 05 月 26 日 (土)
-
0910 起床.
コーヒー.
朝飯.
洗濯.
怠業.
-
昼飯.
頭痛.
寝る.
-
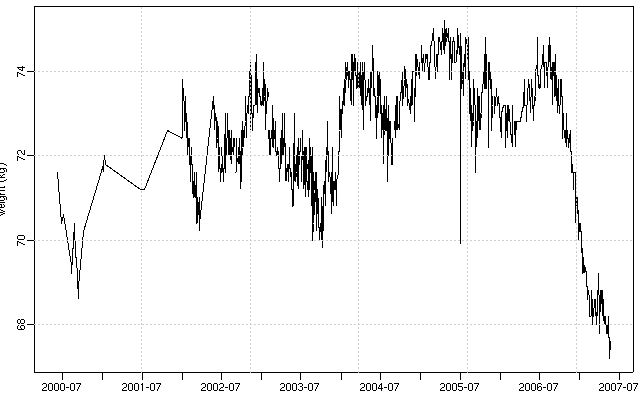
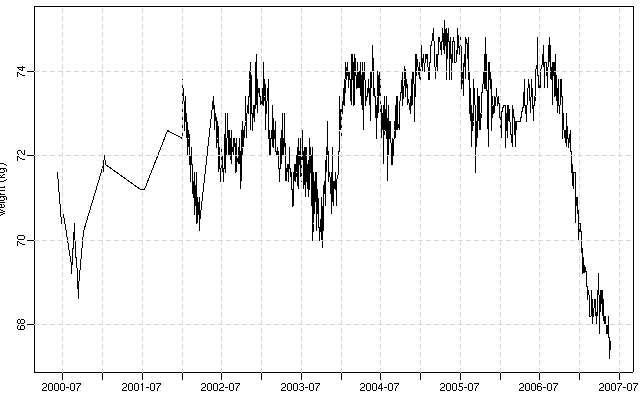
夜になって回復してきたので,
「『ぎょーむ日誌』に記録されてる体重」
を日付とともに抽出する Perl スクリプトつくり,
R
で作図してみる.
データ抽出プログラムが不完全でところどころヘンな値になってしまっているけど
……
-
まあ,
昨年夏以降けっこう減少しました,
と
(cf.
15kg 減量に至らない記録).
-
[今日の運動]
-
腹筋運動 30 ×
3 回.
腕立ふせ 10 ×
3 回.
スクワット 100 回.
-
[今日の食卓]
- 朝 (0950):
米麦 0.5 合.
キュウリ.
トマト.
- 昼 (1330):
イソップの昔ふうバゲット.
ネギ・ジャガイモ・ニンジン・ブロッコリー・ブナピー・
干しエビ・ホタテのクリームシチュー.
- 晩 (1930):
北 14 西 1 グロビュールの豆カレー.
辛さ 3.
2007 年 05 月 27 日 (日)
-
1010 起床.
うう.
コーヒー.
朝飯.
怠業.
-
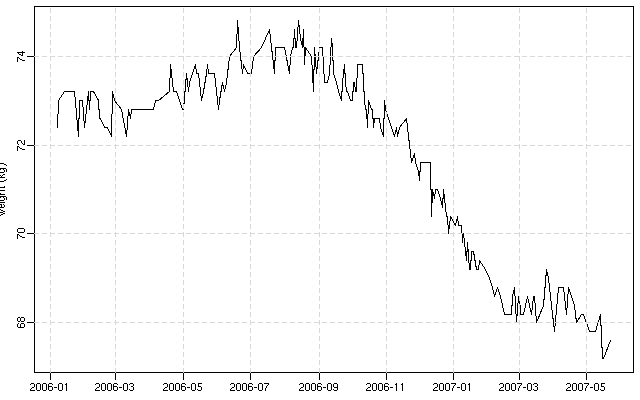
昨日の 6.5 年間体重変化グラフ,
「『ぎょーむ日誌』に記録されてる体重」
を日付とともに抽出する Perl スクリプト
(全データ抽出に必要な時間は 0.5 分ぐらい)
を少しだけ改善して,
図をつくりなおしてみる.
-
これは
2006-01-01
から現在まで.
うーむ,
健康なのか不健康なのか
……
蛇足ながら,
BMI
(body mass index) なるアヤしげな割り算値は
(体重 [kg]) (身長 [m])-2
などと定義されていて,
これが 18-25 ぐらいの値なら太りすぎでもやせすぎでもないんじゃないの,
と憶測するらしい.
現在の体重 68 kg とすると,
68 × 1.77-2 = 21.7
なので,
いたってふつーということになるのか?
-
いつまでもうだうだとしてられないので,
1615 自宅発.
晴やや曇.
1630 研究室着.
-
トドマツ作文修正,
なかなか進まず
……
2015 研究室発.
2040 帰宅.
晩飯.
-
[今日の運動]
-
[今日の食卓]
- 朝昼 (1130):
イソップのクロワッサン.
ヨーグルト.
- 晩 (2140):
米麦 0.8 合.
キャベツ・ピーマン・ニンジン・ネギ・ショウガ・ニンニク・
シイタケ・豚肉の炒めもの.
2007 年 05 月 28 日 (月)
-
0740 起床.
コーヒー.
朝飯.
0940 自宅発.
晴.
0955 研究室着.
-
トドマツ作文修正が進まないので,
とりあえず現時点の修正案を整理して後藤さん・練さんに送信してみる.
下の文章でみょーに漢語と敵国語の表記が多いのは,
カタカナ語とかを減らしたほうが練さんが読みやすいのでは
(ホントか?),
などと考えてしまったため.
私が現在修正しようとしているのは
・階層 Bayes model の説明 (model, 結果, 考察の修正)
・Introduction と discussion における階層 Bayes model の正当化
といったところです.
後者について現在の構想を説明してみます (自分の論点を整理するためにも).
この論文に階層 Bayes model が必要とされる理由はこの観測データに見られ
る pattern を説明するためには,多くの (母樹・倒木に原因のある) random
effects が必要とされるだろう,というところが出発点になるだろうと思いま
す.なぜ random effects があるのか,と考えると,ひとつは
1. 倒木の野外観察によって,近接する同じような場所にある倒木でも
seedling-sapling 密度がまったく異なるように見える (overdispersion)
があげられます.さらに (これは遺伝子解析の結果をみてわかることですが)
cpSSR と nuclear SSR の解析によって詳細な母子関係が判明し,
2. 母樹によって散布距離が異なるように見える
というパターンが判明してしまいました.つまり
疑問: なぜこの調査地の seedling-sapling の空間分布 (pattern, 母子関係
も含む) はこうなってしまうのか?
に解答するために,
- 母樹からの距離,倒木上の面積の効果: fixed effects
- 母樹・倒木の原因不明な効果: random effects
の両者を同時に考慮した階層 Bayes model を構築し,データから posterior
を推定することによって,観測 pattern を説明できるような fixed/random
effects が得られるという話になりそうです.
つまり Introduction では
・調査地現場における野外観察から倒木差が予想される
・高精度 parentage analysis によって母樹差の推定が期待される
をあげてこれら random effects と母子間距離・倒木面積 (こちらは offset
term 的ですが) の fixed effets を分離して推定するには階層 Bayes model
とならざるをえない,といった正当化になりそうです.
いっぽう discussion では,高精度 parentage analysis の「結果」をうけて,
やはり母樹差 random effects を考慮すべきであった,という正当化になりそ
うです.つまり散布距離のばらつきだけであるならば階層 Bayes model は不要
である (単純に母樹ごとに母子間距離を計算すればよい),しかし従来の散布モ
デル (Burczyk 型モデルなど) とこの「母樹によって散布距離が異なる現象」
を対応づけるためには,散布 model (dispersal kernel) に「母樹差」をいれ
た新しいモデルに拡張するほかない,といった内容です.
以上はここまでの私の説明を整理したもので,とくに新しいものはないだろう
と思います.
問題なのは genetic structure を強調する現時点の原稿との対応づけだろうと
思います.しかし,この問題は私の修正作業がいったん終了してからまた議論
したほうが良さそうですね.
補足してみると,
高精度 parentage analysis によって判明した結果を使って
「単純に母樹ごとに母子間距離を計算すればよい」
なる方式は昨年投稿した原稿で使われていたもので,
Burczyk 的な散布モデルと対応つけろというコメントもついていたので,
モデル屋たる私がこの研究プロジェクトによばれた次第
……
なのであった.
ということで,
まずは discussion から修正していくんだけど,
introduction との重複部分を
……
とやってると,
いつのまにか introduction ぢりぢり修正作業になっている,
と.
いつまでもぢりぢり作文ばかりやってる罰というべきか,
永光さんからシウリザクラ再解析用のデータが
……
ちょっと質問メイルを返信.
これはいつもの時間かせぎ,
ではなく.
作文すこしなおしてから昼飯.
6/1 ここ発でまた Pasoh にいく矢澤さんが
R
で調査用データシート
(このあたりで苦闘
→
ぎょーむ日誌 2006-10-23)
の自動生成中.
じつはこの R プログラムは LaTeX file を生成するものであり
……
ゐんどーづ中で何とかしようとすると,
2007-04-22
あたりの作業をくりかえして
WinShell 3.1
一式をインストールすることになる.
TeXインストーラ3
がまた改善されたためなのか,
本日は GhostScript への path 設定 (for dvipdfmx)
が不要だった
(前回は手動で GS インストールしたのがまずかった?).
ただし landscape 用紙設定の PDF file 生成のためには,
PDFTeX
設定を dvipdfmx
とするだけでなく引数設定を
-l "%s.dvi"
としなければならぬ.
トドマツ作文
……
そしてまた Pasoh データシート問題に.
昨年 10 月に私が作った調査用データシート R コード,
なかなか読みづらいものになっていて矢澤さんも投げてしまった
……
というか 50 ha 内の各 1 ha を「適切な長さのペイジ数に分割せよ」という
新しい仕様をくみこもうとして挫折,
ですか.
とりあえず 2 時間ほどかけてプログラム改造.
ゐんどーづ上の (Tinn-R などの) 作業なので隔靴掻痒感あり.
改造を進めるカギは
「難しくなってきたら function() 化」
ということなんだけど,
慣れてないヒトには難しい発想かもしれない.
2045 研究室発.
2100 帰宅.
晩飯の準備.
晩飯.
[今日の運動]
[今日の食卓]
- 朝 (0900):
イソップのバゲット.
キュウリ・レタスのサラダ.
昨晩の残り,
キャベツ・ピーマン・ニンジン・ネギ・ショウガ・ニンニク・
シイタケ・豚肉の炒めもの.
- 昼 (1340):
研究室お茶部屋.
米麦 0.6 合.
もずくスープ.
リンゴ.
- 晩 (2200):
米麦 0.8 合.
タマネギ・ブロッコリー・レタス・ブナピー・豆腐のカレー.
キュウリ・ホウレンソウのサラダ.
2007 年 05 月 29 日 (火)
c <- "#0000ff"
par(col = c, col.axis = c, col.lab = c, col.main = c, fg = c, bg = "#ddddff")
par(mfrow = c(2, 2), mar = c(3, 3, 1, 1), mgp = c(2, 0.5, 0))
for (i in 1:4) plot(rnorm(100), rnorm(100))
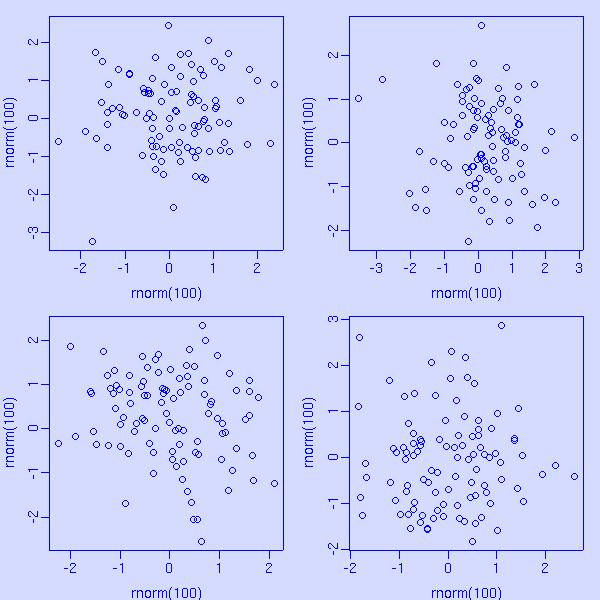
-
しかしその質問院生
(名はここでは明かせない
が Pasoh に行くというので絶好調になっている矢澤さん)
が「何だかヘンです」というので,
拝見しにいくと
……
色指定とは何の関係もない問題だとわかった.
絶好調院生には勝てない.
そして絶好調院生は新しい R わざを見つけるきっかけを提供してくれる.
for(i in XXX) ...
で XXX の部分をふつーの vector とかにしとくと,
どうしても一回は「回ってしまう」わけだが,
> for (i in 1:3) print(i)
[1] 1
[1] 2
[1] 3
> for (i in 1:1) print(i)
[1] 1
> for (i in 1:0) print(i)
[1] 1
[1] 0
しかしながら numeric(0)
とすると,
> for (i in numeric(0)) print(i)
一度も回らない.
蛇足ながら 0 以外の numeric()
は (とうぜんながら) まわる.
> for (i in numeric(3)) print(i)
[1] 0
[1] 0
[1] 0
読みやすくなるかどうかはともかく,
for(i in numeric(0)) ...
わざは何か使えることありそう
(本日は役にたった).
-
多変量正規分布を事前分布とする階層ベイズモデルについてまた検討してしまう
……
はい,
現時点においてこの行為は現実逃避ですね.
-
ということで,
トドマツ作文みなおしのつづき.
-
ひととーりできたというか,
とりあえずの締め切り時刻が近づいてきたので,
make pdf && make save
の rsync でアップロード,
メイルかき,
といつもの手順をふむ.
-
ところで後藤さんから質問されて気づいたのだが,
ゐんどーづ版 Aspell は TeX installer でもインストールされないので
(option を見のがしたか?),
GNU Aspell (Win32 version)
から本体と辞書をダウンロードしてインストールする必要がある.
TeX wiki
にもあるように,
このインストールさえできていれば Winshell は特に設定する必要なく
「スペルチェック」できるようになる.
-
ふーむ,
会計係がアヤしげな個体群調査を企画しているな
……
「建築物における衛生的環境の確保に関する法律」に基づく標記の件
について,下記日程のとおり行うこととなりましたのでお知らせいたし
ます。
調査の際は各部屋に立ち入り,捕獲器の設置を行う場合がありますの
でご協力方お願いいたします。
記
6月 4日 10時〜事前調査
6月 8日 捕獲器設置
6月19日 捕獲器回収
1840 研究室発.
1855 帰宅.
買いもの.
晩飯の準備.
晩飯.
[今日の運動]
[今日の食卓]
- 朝 (0620):
米麦 0.6 合.
タマネギ・ブロッコリー・レタス・ブナピー・豆腐のカレー.
- 昼 (1350):
研究室お茶部屋.
イソップのバゲット.
チーズ.
もずくスープ.
- 晩 (2100):
米麦 0.8 合.
タマネギ・ブロッコリー・ジャガイモ・ソーセイジのスープ.
キャベツ・ニンジン・レタスのサラダ.
2007 年 05 月 30 日 (水)
-
0750 起床.
コーヒー.
いきなりシウリザクラ再計算に没頭してしまう
……
ひととーり再計算が終了したように思えたので,
朝飯.
1010 自宅発.
曇.
1025 研究室着.
-
計算結果を確認して server にアップロードする準備
……
のつもりが web site ディレクトリのちょっとした整理作業になってしまった.
-
研究関連のディレクトリから
なぜかしらこんな古い写真ファイルがみつかったり
……
誤
mv
(← こんなのまでゐきぺであにあるのか!)
の結果なんだろうけど.

[発掘写真: 2 年前の追いコン]
2005-02-16,
ひだまり庭にて,
ということのようで.
先日の
体重グラフ
によれば,
この時期あたりは過去数年間の中で
私の体重の極大期だったみたいだねえ
(現在より +6-7 kg ぐらい)
……
-
ともかくいろいろ整理整頓,
rsync
で同期をとって永光さんに連絡メイル,
と.
-
一件落着 & 今日はなぜか院生密度が低いので,
すばやくお茶部屋で昼飯.
-
専攻科事務室の総務から送りつけられてきたゑくせる (!)
ファイルがおかしいように思えたので,
質問メイル.
しかし大学の事務処理能力って
(かかる方法論的限界のおかげで)
むちゃくちゃ低い
……
10-20 年前と比較すると
「ちょー高性能」
というほかない計算機がてもとにありながら,
やってることは「紙だけ」時代とほとんど変わらん,
という未来を予測できたヒトっているのだろうか?
大学の (少なくとも北大の) 事務系ってぜんぜんだめだめだな.
-
ついでにこういう事務系のヒトたちが電話攻撃を好むのは,
(私にとっては意外だったんだけど)
皆さん作文能力 (とくに説明なんかの) がかなり低めだから,
ってのも一因のような気がする
……
カタにはまった文面のこぴぺ & 微修正は好きなのにね.
-
給料まわりと旅費まわりだけは何とかオンライン化されてる
……
最近は物品購入と実験室の薬品管理も追加されたな.
他大学とほとんど同じシステムが流用できるもの
(つまり給料まわり)
はかなりデキが良い.
他のものは北大かすたまいず & 北大式運用が入ってしまい,
それゆえに使いにくいものになっている.
-
いんふらでは
北大図書館
はかなり良いと思う.
さすが情報処理の専門家というか.
それに対して,
「情報」を名に関してるけど,
北大ねっとはいつまでたっても
へぼいまま
(みかけだけでなく内容・しくみも).
-
蛇足ながらここ専攻科の web まわりもかなり苦闘しているらしい
(先日の会議によれば).
外注して web site 作ってもらったのは良いものの,
内容更新のたびに
どりーむゐーばー
動かさねばならず,
そのたびに大さわぎになるそーで (傍観).
-
そして罰あたりなことばかり書いてたむくいか,
Gmail server がコケてしまった.
時刻は 1400 あたり.
すぐに復旧.
-
しばらく放置していた地衣類繁殖の階層ベイズモデル説明作文に
またとりくんでみる
……
おうぷんおひす
操作はイヤだなぁ.
まあ,
MS わーどよりは精神的苦痛が少ないはず,
だと思うんだけど.
-
えーと,
前回までで岩面の明るさ指数の三次元計算と
階層ベイズモデルの説明はひととーりできてる,
と.
で Results 節にとりくむわけだが
……
そうか図表あれこれの準備が必要だったわけだな.
-
ということで,
R & LaTeX で楽しく作図.
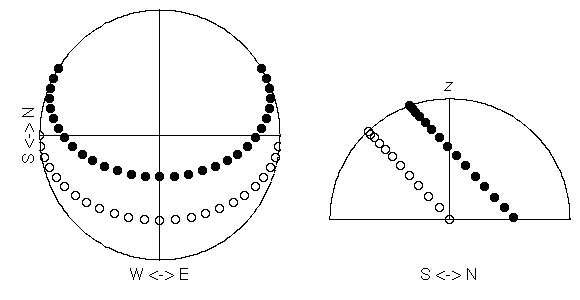
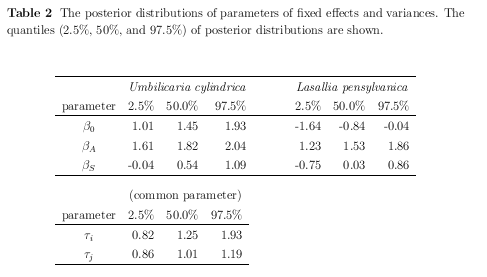
Fig.4 (a), (b)
それぞれ
Lasallia pensylvanica
(オオイワブスマ)
と
Umbilicaria cylindrica
(タカネコゲノリ)
の「好む明るさ」 (岩面の方位・傾斜できまる)
における子器 (apothecia) 密度.
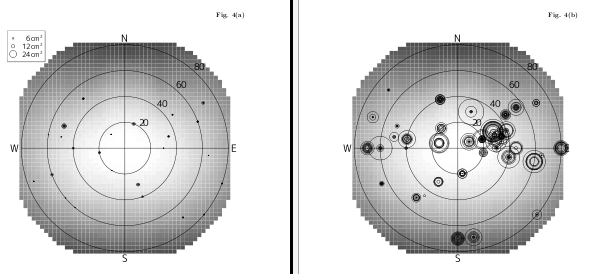
-
Table も R & LaTeX で楽しく,
と.
-
うーむ,
MCMC な図も追加したほうがよいかも,
と
……
思いつつ撤退.
1930 研究室発.
1945 帰宅.
ばてた.
体重 67.8kg.
晩飯.
-
[今日の運動]
-
[今日の食卓]
- 朝 (0930):
米麦 0.6 合.
タマネギ・ブロッコリー・ジャガイモ・ソーセイジのスープ.
キャベツ・ニンジン・レタスのサラダ.
- 昼 (1250):
研究室お茶部屋.
米麦 0.6 合.
タマネギ・ブロッコリー・レタス・ブナピー・豆腐のカレー.
- 晩 (2200):
米麦 0.7 合.
ジャガイモ・タマネギ・マイタケ・鶏肉の煮物.
キャベツ・ニンジン・タマネギ・トマトのサラダ.
2007 年 05 月 31 日 (木)
-
0730 起床.
コーヒー.
朝飯.
0855 自宅発.
晴.
0910 研究室着.
-
セミナーの予習.
-
1030 より
研究室セミナー,
本日は宮田さんでシュートあれこれの樹高依存性論文の紹介.
- Ishii, H. et al. (2002) Variation in specific needle area of old-growth Douglas-fir in relation to needle age, within-crown position and epicormic shoot production. Tree Physiology. 22: 31-40
Variation in specific needle area (SNA; cm2 projected
fresh needle area g-1 oven-dried needle weight) was investigated
in relation to needle age, within-crown position and
epicormic shoot production in 450-year-old Douglas-fir
(Pseudotsuga menziesii Mirb. (Franco) var. menziesii) trees.
Specific needle area decreased with increasing needle age. The
magnitude and rate of change in SNA with needle age were
greatest for lower-crown branches, and decreased toward the
middle- and upper-crown branches. For all branches, there was
no difference between regular and epicormic shoots in the relationship
between SNA and needle age.
...
- Koch, G.W. (2004) The limits to tree height. Nature. 428;851-854.
Trees grow tall where resource are abundant, stresses are minor,
and competition for light places a premium on height growth.
The heigh to which trees can grow and the biophysical determinants
of maximum height are poorly understood. Some models
predict heights of up to 120 m in the absence of mechanical
damage, but there are historical accounts of taller trees.
Current hypotheses of hight limitation focus on increasing
water transport constraints in taller trees and the resulting
reductions in leaf photosynthesis. We studied redwoods
(Sequoia sempervirens), including the tallest known tree on
Earth (112.7 m), in wet temperate forests of northern California.
Our regression analyses of height gradients in leaf functional
characteristics estimate a maximum tree height of 122-130 m
barring mechanical damage, similar to the tallest recorded trees
of the past. As trees grow taller, increasing leaf water stress
due to gravity and path length resitance may ultimately limit leaf
expansion and photosynthesis for further height growth, even
with ample soil moisture.
- Matsuzaki, J. et al. (2005) Shoots grafted into the upper crowns of tall Japanese cedar (Cryptomeria japonica D. Don) show foliar gas exchange characteristics similar to those of intact shoots. Trees. 19: 198-203
The lower foliar photosynthetic rates seen in
shoots in the upper crowns of tall trees than those in
seedlings could be caused by extrinsic factors, such as
hydraulic conductance, for shoots or by irreversible
intrinsic change in the meristems during tree development.
To clarify which is most significant, we compared foliar
gas exchange charcteristics and water relations among
scions of Japanese cedar (Cryptomeria japonica D. Don)
grafted into the upper crowns of tall trees, rooted cuttings
developed from scions of the same clones, and intact
shoot in the upper crowns of the tall trees.
...
- McDowell, N. et al. (2002) The relationships between tree height and leaf area: sapwood area ratio. Oecologia. 132: 12-20
The leaf area to sapwood are ratio (Al:As) of
trees has been hypothesized to decrease as trees become
older and taller. Theory suggests that
Al:As
must decrease to maintaion leaf-specific hydraulic sufficiency as path
length, gravity, and tourtuosity constaratin whole-plant hydraulic
conductance.
...
- Nabeshima, E. and Hiura, T. (2004) Size dependency of photosynthetic water- and nitrogen-use efficiency and hydraulic limitation in Acer mono. Tree Physiology. 24: 745-752.
We examined open-growth Acer mono Maxim.
trees of different sizes to test the hypothesis that (1) hydraulic
limitation increases with tree size, thereby reducing photosynthesis,
and (2) photosynthetic water- and nitrogen-use efficiencies
chnage with tree size. Maximum net assimilation rate per
unit dry mass was significantly lower in large trees thant in
small trees, whereas leaf nitrogen concentration increased with
tree size. As a consequence, photosynthetic nitrogen-use efficiency
decreased with increase in tree size, partly as a result of reduced
of stomatal conductance. Neither root-to-leaf hydraulic conductance
nor minimum leaf water potential changed with tree size.
- Osada, N. et al. (2004) Changes in crown development patterns and current-year shoot structure with light environment and tree height in Fagus crenata (Fagaceae). Ammerican Journal of Botany. 91: 1981-1989
The relative effects of light and tree height on the architecture of leader crowns
(i.e. the leading section of the main trunk, 100 cm in length) and current-year shoots
for canopy species, Fagus crenata, occupying both the ridge top and the valley
bottom in a cool-temperate forest in Japan were investigated. For leader crowns,
the number of current-year shoots and leaves increased with increasing tree height,
such that the relative allocation to stems increased with increasing tree height.
Furthermore, stem structures within current-year shoots also changed with height,
such that taller trees produced thicker and shorter stems of the same volume.
...
まあ,
いろいろな論文あげられたわけだが,
宮田さんまわりの苫小牧樹高研究と
(上にあげられた論文の多くが着目している)
水分制約が樹高を制限しているといったハナシはまだうまく整理できてない.
で,
本年は分枝数というか当年枝姉妹集団も調べる,
と
……
まあ,
昨年の当年枝サンプルの食害 (虫くい) 状況も最近しらべてみたいだから,
そちらの影響を考慮して推定計算とかやりなおすほうが先かな.
-
上の中で Koch et al. (2004) は樹高 100 m 超のてっぺんとかで
水ストレスや光合成速度を測定していて,
さらにとってきたシュートを実験室で「いけばな」状態にして
また光合成速度とか測っている.
その結果,
-
面積あたり光合成速度
(今までよく考えてなかったけどこれも割算値の世界だな)
は樹高依存性がない
-
しかし重量あたり光合成速度だと樹高依存性があり,
シュート採取樹高とともに光合成速度は落ちるらしい
(原因はよくわからん,とのこと)
-
さらに上記 Matsuzaki et al. (2005) はスギの「つぎ木」実験していて,
5-6 m ぐらいのところからとったシュートを 25-30 m ぐらいにつないで,
(1.5 年後に) その光合成速度などを測定している.
するともともとその樹高にあったシュートと光合成速度は変わらなかった,
と.
-
うーむ,
これらの研究では測定条件
(切ってきてすぐに測定 vs 時間かけてつぎ木)
が異なっているので,
単純には比較できんな.
今年 3 月の生態学会松山大会の
枝自由集会
で石井さん (神戸大) がこういった「つぎ木」実験の多様な結果あれこれ
に言及しておられたな
……
まあ,
今日のセミナーではここまでふみこんだ議論はなかったんだけど.
-
ということで,
セミナー終了後もこのへんの論文など調べてしまった.
-
1300 すぎにお茶部屋で昼飯.
小林君と北海道ブナ林遺伝子構造雑談.
うーむ,
私からみるとけっこうナゾの構造
(もしくはすごーく単純に形成された構造),
のような
……
-
大雪山系稜線上の地衣類繁殖の階層ベイズモデル論文の図の準備,
MCMC 図を作ってみる
……
うーん,
こういうのはどうなんだろう?
これまた電気鼠完全無用の R & LaTeX 複合わざ.
R プログラミングもふくめて
作業時間は 2 時間ほどかな?
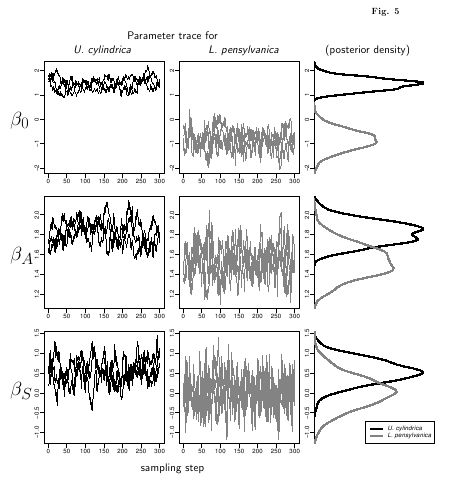
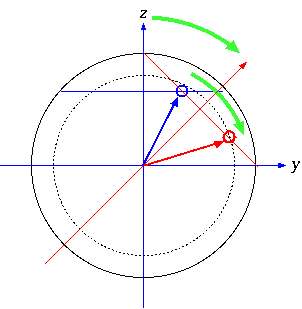
\setlength{\unitlength}{0.1mm}
\begin{picture}(1900,2000) %%% 1800 + 100, 1800 + 2000
\Huge
\put(100,50){\includegraphics[width=180mm]{mcmc.eps}}
\put(0,1550){$\beta_0$}
\put(0,950){$\beta_A$}
\put(0,350){$\beta_S$}
\large\sffamily
\put(520,1930){Parameter trace for}
\put(280,1870){\textit{U. cylindrica}}
\put(880,1870){\textit{L. pensylvanica}}
\put(1400,1870){(posterior density)}
\put(580,10){sampling step}
\end{picture}
-
今まで作ってきた図表のこまかい修正作業などなど.
うーむ,
あちこちに修正すべき点が
……
-
最後はおうぷんおひすで figure legends を少し書いて,
撤退.
1845 研究室発.
1900 帰宅.
晩飯.
-
[今日の運動]
-
腹筋運動 30 ×
3 回.
腕立ふせ 10 ×
3 回.
-
[今日の食卓]
- 朝 (0800):
クラッカー.
キャベツ・ニンジン・タマネギ・トマトのサラダ.
- 昼 (1320):
研究室お茶部屋.
食パン.
チーズ.
- 晩 (2040):
米麦 0.8 合.
タマネギ・ネギ・ピーマン・シイタケ・卵の炒めもの.