ぎょーむ日誌 2007-05-31
2007 年 05 月 31 日 (木)
-
0730 起床.
コーヒー.
朝飯.
0855 自宅発.
晴.
0910 研究室着.
-
セミナーの予習.
-
1030 より
研究室セミナー,
本日は宮田さんでシュートあれこれの樹高依存性論文の紹介.
- Ishii, H. et al. (2002) Variation in specific needle area of old-growth Douglas-fir in relation to needle age, within-crown position and epicormic shoot production. Tree Physiology. 22: 31-40
Variation in specific needle area (SNA; cm2 projected
fresh needle area g-1 oven-dried needle weight) was investigated
in relation to needle age, within-crown position and
epicormic shoot production in 450-year-old Douglas-fir
(Pseudotsuga menziesii Mirb. (Franco) var. menziesii) trees.
Specific needle area decreased with increasing needle age. The
magnitude and rate of change in SNA with needle age were
greatest for lower-crown branches, and decreased toward the
middle- and upper-crown branches. For all branches, there was
no difference between regular and epicormic shoots in the relationship
between SNA and needle age.
...
- Koch, G.W. (2004) The limits to tree height. Nature. 428;851-854.
Trees grow tall where resource are abundant, stresses are minor,
and competition for light places a premium on height growth.
The heigh to which trees can grow and the biophysical determinants
of maximum height are poorly understood. Some models
predict heights of up to 120 m in the absence of mechanical
damage, but there are historical accounts of taller trees.
Current hypotheses of hight limitation focus on increasing
water transport constraints in taller trees and the resulting
reductions in leaf photosynthesis. We studied redwoods
(Sequoia sempervirens), including the tallest known tree on
Earth (112.7 m), in wet temperate forests of northern California.
Our regression analyses of height gradients in leaf functional
characteristics estimate a maximum tree height of 122-130 m
barring mechanical damage, similar to the tallest recorded trees
of the past. As trees grow taller, increasing leaf water stress
due to gravity and path length resitance may ultimately limit leaf
expansion and photosynthesis for further height growth, even
with ample soil moisture.
- Matsuzaki, J. et al. (2005) Shoots grafted into the upper crowns of tall Japanese cedar (Cryptomeria japonica D. Don) show foliar gas exchange characteristics similar to those of intact shoots. Trees. 19: 198-203
The lower foliar photosynthetic rates seen in
shoots in the upper crowns of tall trees than those in
seedlings could be caused by extrinsic factors, such as
hydraulic conductance, for shoots or by irreversible
intrinsic change in the meristems during tree development.
To clarify which is most significant, we compared foliar
gas exchange charcteristics and water relations among
scions of Japanese cedar (Cryptomeria japonica D. Don)
grafted into the upper crowns of tall trees, rooted cuttings
developed from scions of the same clones, and intact
shoot in the upper crowns of the tall trees.
...
- McDowell, N. et al. (2002) The relationships between tree height and leaf area: sapwood area ratio. Oecologia. 132: 12-20
The leaf area to sapwood are ratio (Al:As) of
trees has been hypothesized to decrease as trees become
older and taller. Theory suggests that
Al:As
must decrease to maintaion leaf-specific hydraulic sufficiency as path
length, gravity, and tourtuosity constaratin whole-plant hydraulic
conductance.
...
- Nabeshima, E. and Hiura, T. (2004) Size dependency of photosynthetic water- and nitrogen-use efficiency and hydraulic limitation in Acer mono. Tree Physiology. 24: 745-752.
We examined open-growth Acer mono Maxim.
trees of different sizes to test the hypothesis that (1) hydraulic
limitation increases with tree size, thereby reducing photosynthesis,
and (2) photosynthetic water- and nitrogen-use efficiencies
chnage with tree size. Maximum net assimilation rate per
unit dry mass was significantly lower in large trees thant in
small trees, whereas leaf nitrogen concentration increased with
tree size. As a consequence, photosynthetic nitrogen-use efficiency
decreased with increase in tree size, partly as a result of reduced
of stomatal conductance. Neither root-to-leaf hydraulic conductance
nor minimum leaf water potential changed with tree size.
- Osada, N. et al. (2004) Changes in crown development patterns and current-year shoot structure with light environment and tree height in Fagus crenata (Fagaceae). Ammerican Journal of Botany. 91: 1981-1989
The relative effects of light and tree height on the architecture of leader crowns
(i.e. the leading section of the main trunk, 100 cm in length) and current-year shoots
for canopy species, Fagus crenata, occupying both the ridge top and the valley
bottom in a cool-temperate forest in Japan were investigated. For leader crowns,
the number of current-year shoots and leaves increased with increasing tree height,
such that the relative allocation to stems increased with increasing tree height.
Furthermore, stem structures within current-year shoots also changed with height,
such that taller trees produced thicker and shorter stems of the same volume.
...
まあ,
いろいろな論文あげられたわけだが,
宮田さんまわりの苫小牧樹高研究と
(上にあげられた論文の多くが着目している)
水分制約が樹高を制限しているといったハナシはまだうまく整理できてない.
で,
本年は分枝数というか当年枝姉妹集団も調べる,
と
……
まあ,
昨年の当年枝サンプルの食害 (虫くい) 状況も最近しらべてみたいだから,
そちらの影響を考慮して推定計算とかやりなおすほうが先かな.
-
上の中で Koch et al. (2004) は樹高 100 m 超のてっぺんとかで
水ストレスや光合成速度を測定していて,
さらにとってきたシュートを実験室で「いけばな」状態にして
また光合成速度とか測っている.
その結果,
-
面積あたり光合成速度
(今までよく考えてなかったけどこれも割算値の世界だな)
は樹高依存性がない
-
しかし重量あたり光合成速度だと樹高依存性があり,
シュート採取樹高とともに光合成速度は落ちるらしい
(原因はよくわからん,とのこと)
-
さらに上記 Matsuzaki et al. (2005) はスギの「つぎ木」実験していて,
5-6 m ぐらいのところからとったシュートを 25-30 m ぐらいにつないで,
(1.5 年後に) その光合成速度などを測定している.
するともともとその樹高にあったシュートと光合成速度は変わらなかった,
と.
-
うーむ,
これらの研究では測定条件
(切ってきてすぐに測定 vs 時間かけてつぎ木)
が異なっているので,
単純には比較できんな.
今年 3 月の生態学会松山大会の
枝自由集会
で石井さん (神戸大) がこういった「つぎ木」実験の多様な結果あれこれ
に言及しておられたな
……
まあ,
今日のセミナーではここまでふみこんだ議論はなかったんだけど.
-
ということで,
セミナー終了後もこのへんの論文など調べてしまった.
-
1300 すぎにお茶部屋で昼飯.
小林君と北海道ブナ林遺伝子構造雑談.
うーむ,
私からみるとけっこうナゾの構造
(もしくはすごーく単純に形成された構造),
のような
……
-
大雪山系稜線上の地衣類繁殖の階層ベイズモデル論文の図の準備,
MCMC 図を作ってみる
……
うーん,
こういうのはどうなんだろう?
これまた電気鼠完全無用の R & LaTeX 複合わざ.
R プログラミングもふくめて
作業時間は 2 時間ほどかな?
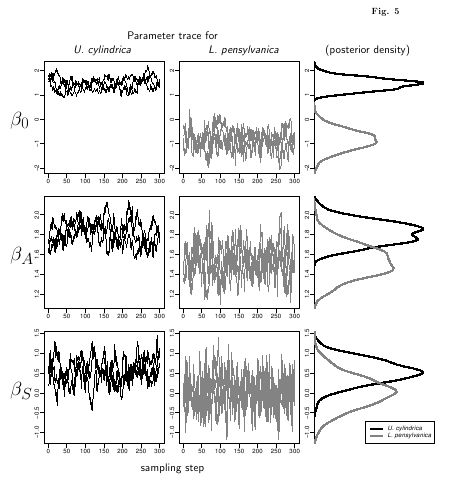
\setlength{\unitlength}{0.1mm}
\begin{picture}(1900,2000) %%% 1800 + 100, 1800 + 2000
\Huge
\put(100,50){\includegraphics[width=180mm]{mcmc.eps}}
\put(0,1550){$\beta_0$}
\put(0,950){$\beta_A$}
\put(0,350){$\beta_S$}
\large\sffamily
\put(520,1930){Parameter trace for}
\put(280,1870){\textit{U. cylindrica}}
\put(880,1870){\textit{L. pensylvanica}}
\put(1400,1870){(posterior density)}
\put(580,10){sampling step}
\end{picture}
-
今まで作ってきた図表のこまかい修正作業などなど.
うーむ,
あちこちに修正すべき点が
……
-
最後はおうぷんおひすで figure legends を少し書いて,
撤退.
1845 研究室発.
1900 帰宅.
晩飯.
-
[今日の運動]
-
腹筋運動 30 ×
3 回.
腕立ふせ 10 ×
3 回.
-
[今日の食卓]
- 朝 (0800):
クラッカー.
キャベツ・ニンジン・タマネギ・トマトのサラダ.
- 昼 (1320):
研究室お茶部屋.
食パン.
チーズ.
- 晩 (2040):
米麦 0.8 合.
タマネギ・ネギ・ピーマン・シイタケ・卵の炒めもの.