ぎょーむ日誌 2007-06-(11-20)
2007 年 06 月 11 日 (月)
-
0810 起床.
コーヒー.
朝飯.
0925 自宅発.
晴.
0940 研究室着.
-
またアリデータにとりくむ.
昨日の脱ゑくせるデータさるべいじ作業,
文字コードがらみで一ヶ所まちがいある,
とわかった.
修正.
「逆襲データ除去しました」
表示をちゃんとやらねば,
と表示させててきづいた.
ケガれ言語 Perl まわりの
use encoding ...;
と
use Encode;
もぢりぢりと挙動が理解できてきた.
$ make
./conv.pl allinone.txt > data.csv
# reading keys.txt ...
# NOTICE: removing COUNTER ATTACK item !!! (in line 189)
# (DATA) 9 a "a , b→星置(7sec)" a a a
# -----> 9 a "a a a a
(R を起動)
> d <- read.csv("data.csv")
> summary(as.factor(apply(d[,7:ncol(d)], 1, sum))) # とうぜん昨日とはちがう……
1 2 3
1598 51 1
R
を使った「表の集計」ってのはじつにお手軽なんだけど
……
まだ慣れてないヒトたちには下のような
apply 系関数
(sapply(), apply(), tapply())
の連発わざはちょっとイヤなかんぢかもしれない
(dataframe d の構造は今のところ
こうなってる).
> v.key <- apply(d, 1, function(x) paste(x["tr30m"], x["colony"], x["home"]))
> st <- sapply(7:ncol(d), function(i) tapply(d[, i], v.key, sum))
> colnames(st) <- colnames(d)[7:ncol(d)]
> st
aggdisplay allogrooming antennation biting ignorance others trophallaxis
0 hoshioki atsuta 1 0 71 23 51 5 0
0 hoshioki hoshioki 0 0 53 0 95 2 1
0 hoshioki ishikari 2 0 93 4 49 6 1
0 hoshioki jozankei 2 1 101 20 31 0 0
0 hoshioki oshoro 1 0 107 23 17 5 0
0 hoshioki shinkawa 0 0 90 3 57 5 0
0 hoshioki tarukawa 2 0 79 7 60 10 1
1 atsuta atsuta 0 0 91 4 57 1 0
1 hoshioki hoshioki 0 0 83 1 66 1 0
1 jozankei jozankei 1 0 92 35 40 0 0
1 oshoro oshoro 0 0 91 2 53 6 0
「処理のタグ」である v.key
をもうちょっとひねくって書きなおすと,
こうなる.
やっぱり,
ゑくせる上で「目と手で」集計された値とはびみょーに数が異なります,
と.
aggd allg antn btng ignr othr trph
0_0_0_hoshioki_hoshioki 0 0 53 0 95 2 1
0_1_0_hoshioki_ishikari 2 0 93 4 49 6 1
0_1_0_hoshioki_shinkawa 0 0 90 3 57 5 0
0_1_0_hoshioki_tarukawa 2 0 79 7 60 10 1
0_1_1_hoshioki_atsuta 1 0 71 23 51 5 0
0_1_1_hoshioki_jozankei 2 1 101 20 31 0 0
0_1_1_hoshioki_oshoro 1 0 107 23 17 5 0
1_0_0_hoshioki_hoshioki 0 0 83 1 66 1 0
1_0_1_atsuta_atsuta 0 0 91 4 57 1 0
1_0_1_jozankei_jozankei 1 0 92 35 40 0 0
1_0_1_oshoro_oshoro 0 0 91 2 53 6 0
上の列名 (colnames(d)) を
「自動圧縮」しているのは
関数 abbreviate()
である.
> abbreviate(colnames(d)[7:ncol(d)])
aggdisplay allogrooming antennation biting
"aggd" "allg" "antn" "btng"
ignorance others trophallaxis
"ignr" "othr" "trph"
行動を「まとめる」ってのはちょっとイヤなんだけど
……
たとえばこうしてみたらどうだろうか?
# grouping behavior types ...
d$attack <- d$aggdisplay + d$biting
d$nonhostile <- d$allogrooming + d$others + d$trophallaxis
v.behavior <- c("nonhostile", "ignorance", "antennation", "attack")
-
d$attack:
威嚇される・噛みつかれる,
summary(as.factor(d$attack))
で調べると 330 個体 (1650 回の観察) の全行動みると,
攻撃されたのは 129 回で,
そのうち「威嚇されて噛みつかれた」は 2 回
-
d$nonhostile:
「毛づくろい」される・何か食べものわたされる
(d$trophallaxis)
・わたされてるみたいだけど良く見えなかった
(d$others),
summary(as.factor(d$nonhostile))
でこれは 45 回で重複行動ナシ
-
「敵対性」は
v.behavior
内の順序のごとく上がっているのか? :
ignorance
には「触角で拉致られアリが触られたんだけど,
すぐにチェック終了」
だけでなく
「触られた瞬間に拉致られアリが全力で逃げ出した」
も含まれ,
antennation
は触角でしつこくチェックされてる状況
そして一回の観察で複数の行動が観察されることもあるから,
合計は 1650 回よりちょっと多くなってしまう (+ 53 回),
と.
> sapply(v.behavior, function(k) sum(d[, k]))
nonhostile ignorance antennation attack
45 576 951 131
> sum(d[, v.behavior])
[1] 1703
行動の重複はどうなってるのか,
これを二重 sapply() わざで調べてみると,
> sapply(v.behavior, function(i)
+ sapply(v.behavior, function(j) sum((d[,i] > 0) * (d[,j] > 0))))
nonhostile ignorance antennation attack
nonhostile 45 0 21 0
ignorance 0 576 0 0
antennation 21 0 951 30
attack 0 0 30 129
antennation
されたあげくに仲良くされてる事例が 21,
などと解釈してよいのかしらん?
ともあれこういう grouping やってしまうと
antennation
だけが他の行動
(nonhostile, attack)
とかぶってるわけで
……
とするとですよ,
ここで「行動連鎖」
みたいなものがあると考えたほうが説明しやすいのではないかな?
まず最初に地元アリが拉致られアリを「触角でチェックしてみる」
という行動があって
(そういう接触があった場合のみを観察・記録しているから,
拉致られ → 地元も含む?),
-
瞬間的に判断をくだす
(699 回)
→ ……
-
nonhostile: すぐに仲良しになる
(24 回)
-
ignorance: すぐに放置,
ただし拉致られアリのすばやい逃走も含む
(576 回)
-
attack:
「こいつは敵だ!」
といきなり攻撃
(99 回)
-
antennation:
時間をかけたしつこいチェック
(951 回)
→ ……
-
antennation + nonhostile: やっぱり仲良しになる
(21 回)
-
antennation のみ:
まあ OK でしょうと放置,
あるいは拉致られアリがスキをみて逃げだす
(900 回)
-
antennation + attack:
「やはり敵だったか!」
とおもむろに攻撃
(30 回)
このように整理してよいのであれば,
モデリングはより簡単になりそうだな.
地元アリが拉致られアリに対して
antennation
なんかをやると「攻撃していいのかな? ダメかな?」
と猜疑にとりつかれたような状況になってしまって,
攻撃する確率が減る,
ということなのかしらん?
ちなみに「攻撃しちゃいかん相手
(拉致られアリが同巣出身の場合)
でも攻撃した回数」
は「瞬間的な判断」やると 99 回中の 22 回で,
「しつこくチェック」後には 30 回中の 21 回,
と.
わけわからんね.
アリは時間かけて考えるのが苦手なのか?
> dsub <- d[d$identical.colony == 1,]
> tapply(dsub$attack, dsub$antennation, sum)
0 1
22 21
というか,
上の st なる dataframe みればわかるように,
この 43 回の friendly fire のうちじつに 35 回は,
「孤立したコロニー」ってことになってる
定山渓 (正確には八剣山?)
vs 定山渓 (巣穴から 30 m 離れたところで採取) の場合になってるね.
まわりが敵ばかりだとアリも神経質・凶暴になるのかしらん?
昼飯とるべくいったん帰宅.
1310 研究室発.
暑い.
1400 研究室もどる.
例のよって,
「全データを見ることのできる図」
を作ってみる.
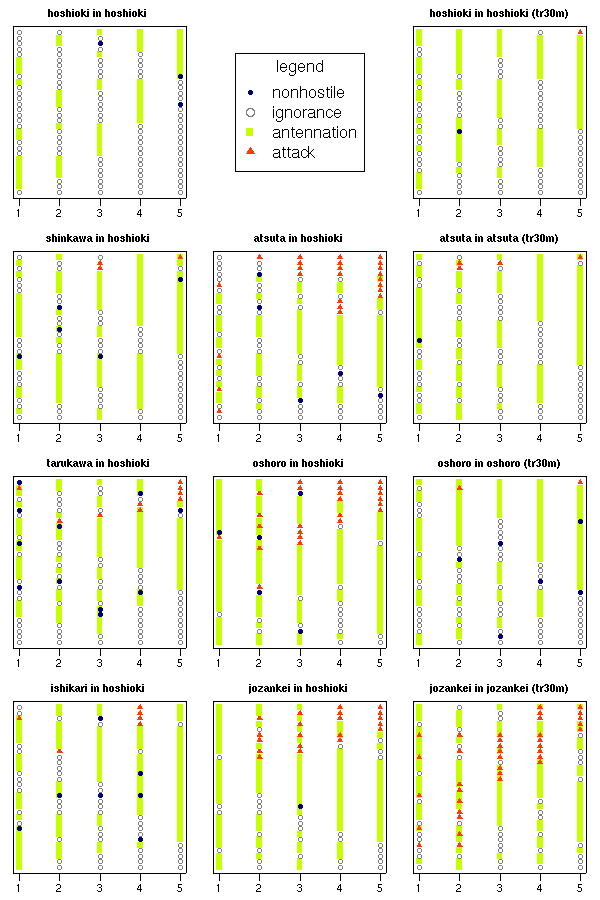
ある処理のタテ軸は 30 個体ぶん,
横軸はその個体の 5 回の「接触」.
「見やすい順」
に order()
するワザをつかっているので,
処理ごとにどういうパターンで攻撃されているかわかる.
外国旅行に伴う航空運賃のクラスについて
平成19年4月の旅費規則・規程の改正に伴い,外国旅行で利用できる航空
運賃のクラスが変更となっていますので,旅券の手配等に留意願います。
1.エコノミークラスで購入しなければならない地域(役員除く)
本邦と次の地域との間の航空旅行
インドネシア,ベトナム,カンボジア,北朝鮮,シンガポール,タイ,大韓民国,
台湾,中華人民共和国,東チモール,フィリピン,ブルネイ,マレイシア,ミャン
マー,モンゴル, ラオス,ハワイ諸島,グアム,ウラジオストク,ハバロフスク
及びユジノサハリンスク
*改正前は,助教授相当以上がビジネスクラスを利用できましたが,改正後
はエコノミークラスに変更になりました。
2.ビジネスクラスを利用できる地域(役員除く)
(1)上記の地域以外の場合において,一の旅行区間における所要航空時
間が8 時間以上の航空旅行(中近東・欧州・北米等)
(2)上記の地域を経由して,目的地までの所要航空時間が8 時間以上の航
空旅行(成田→マレイシア経由→イタリア 等)
3.その他
エコノミークラスしか利用できない外国旅行で,ビジネスクラスを利用し
た場合は,差額分がわかる書類を添付願います。
[今日の運動]
[今日の食卓]
- 朝 (0830):
米麦 0.7 合.
ネギ・卵の炒めもの.
コマツナのゴマあえ.
キャベツ・ニンジン・キュウリのサラダ.
- 昼 (1330):
米麦 0.7 合.
ピーマン・卵炒飯.
キャベツ・ニンジン・キュウリのサラダ.
- 晩 (2030):
ごはんやデンスケ.
日替わり弁当,
塩サバ.
2007 年 06 月 12 日 (火)
-
0830 起床.
コーヒー.
朝飯.
1000 自宅発.
晴.
1015 研究室着.
-
昨日の計算結果でた
……
収束もうちょい.
まだモデルの改良が必要だ.
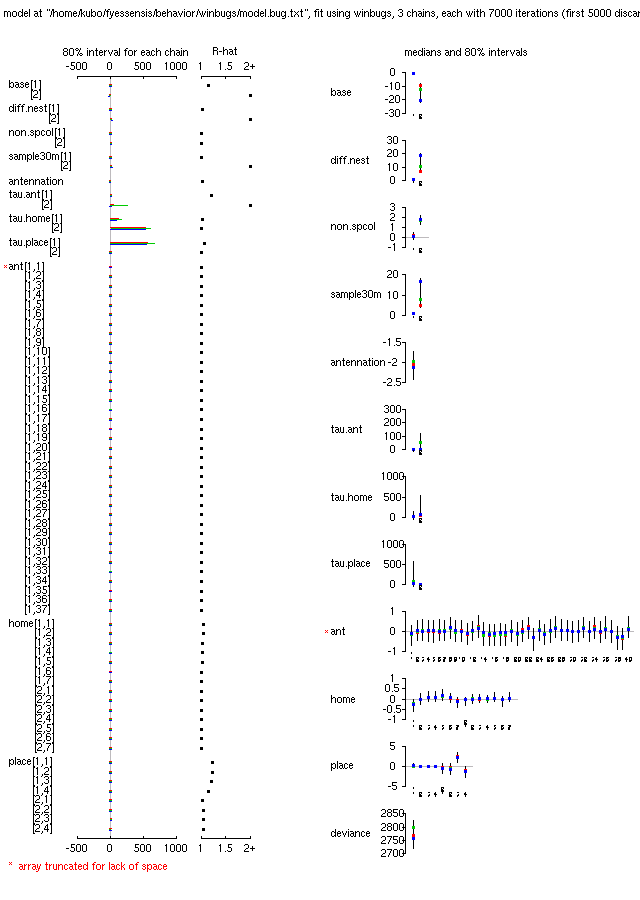
-
こうなる原因はおそらく定山渓アリの異様な凶暴さ (攻撃頻度がすごく高い),
と何か関係ありそう.
-
定山渓アリが home で勝負する実験は 1 set しかない
-
しかもそれは巣から 30 m 離れた「同巣」
(とされてる)
定山渓アリに対するものしかない
つまり「定山渓 home」 & 「定山渓 vs 定山渓バトル」
& 「30 m 離れたところから sampling」
が交絡している特殊な状況で,
「攻撃」が異様な頻度で発生している,
とういことで攻撃まわりのパラメーターが収束してくれません,
と.
-
攻撃に関するモデルをもうちょい簡素化して再計算を命じる.
2000 MCMC step ぐらいで収束してくれないかしらん
……
よし,
収束は格段に改善された.
-
なぜか「アリの『個体差』の分散」パラメーターの収束だけが悪い.
個体差が多いので収束に時間かかる,
ということかしらん?
とりあえず初期値の与えかたで何とかならんかと試行錯誤してみる.
なぜか antennation 受けやすさの
「個体差」
がときどき小さくなりすぎるんだよね
……
-
メイル書いたり要旨とか書いたりしながらモデルの改良.
アリ「個体差」は一種類でよさそう,
とわかってきた.
というのも触角によるしつこいチェック (antennation)
受けたアリは攻撃されることはまれであるからだ
……
攻撃される場合は「こいつは敵だ!」と瞬間的に
判断されて攻撃される確率がかなり高い.
-
といろいろ工夫して 2000 MCMC step (計算時間約 600 秒)
でだいたい収束するようになった
……
と思ったけど,
詳細にみてみるともうちょっとだけ計算時間をのばしたほうがよさそう,
ということで 3000 MCMC step にしてみる.
-
アリ研究会の要旨を
でっちあげて
提出してしまう
……
こういうでっちあげは自分のクビをしめるようなもんだけど,
まあ緊急避難的に.
少なくとも行動解析の階層ベイズモデルについては
エラそーな発表ができるだろう.
-
拉致られアリ計算,
330 個体のアリ「個体差」の幅をさらに制約すると,
計算がうまく収束するようになった.
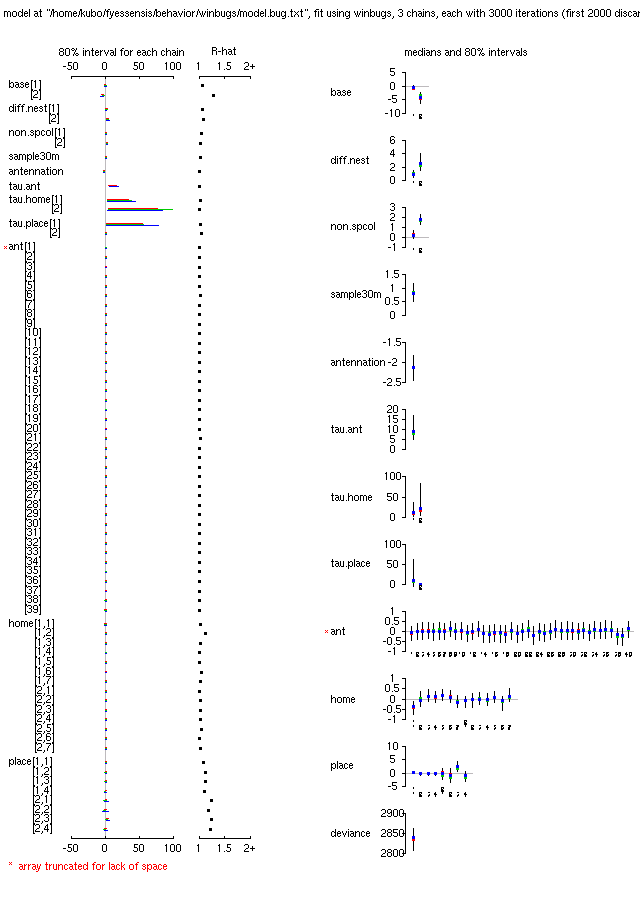
-
これで収束するとなると,
少し欲がでてしまってもうちょいモデルを複雑化したくなるね.
make save
しておいてから,
攻撃モデルに「30 m 遠方効果」いれてみる.
はてさて,
どうなることやら
……
と計算させておいて昼飯.
-
今日は札幌のほうが那覇より気温たかかったそーで
……
明日も暑いみたいだ.
この季節,
北海道は寒暖の差が極端だねえ.
-
……
やはり複雑化モデルはダメ,
とわかった.
まあ現状のままで十分に現象は説明できてるような.
-
「(実験対象である) 拉致られアリがだんだん『疲れて』くる」
効果はどうだろう?
といれてみてまた試しの計算.
その間,
トドマツメイル書き.
-
1000 秒ちょいで計算終了.
ふーむ,
antennation に関しては「疲れ」
(
trial)
効果はないけど,
攻撃を受けることに関してはやっぱりはっきりした傾向あるみたいね.
「アリのココロの中」
モデリングする隠れ状態「負け犬」効果は今回は導入できそうにないけど,
「個体差」 + 「疲れ」で説明できたことにしようかな
……
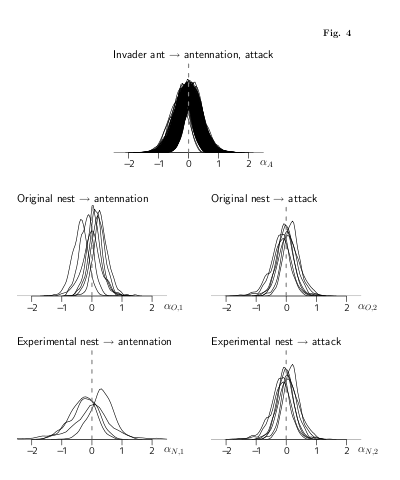
plot(ant.bugs)
は下の図のとーりで,
Table 形式の
print(ant.bugs, digits.summary = 2)
はこちらに.
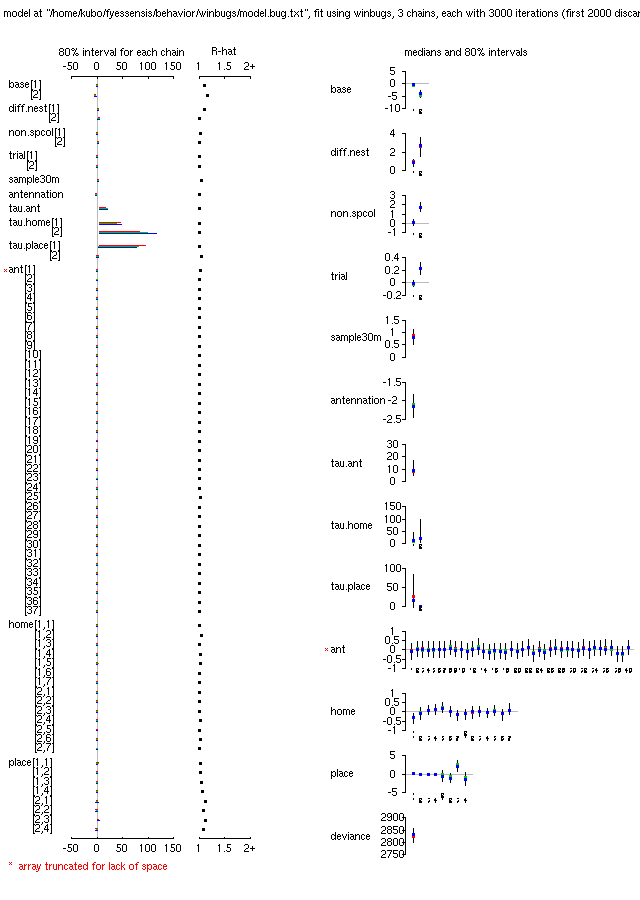
-
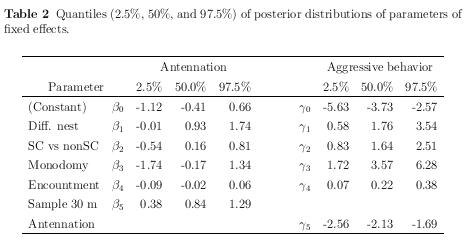
ふーむ,
antennation または攻撃の確率のオッズ (リスク) がいろいろな要因の
係数 (パラメーター) で変わるわけだが,
これらパラメーターの事後分布 (の,そーですね,今回は平均値)
みていくと,
いろいろと興味ぶかい
……
-
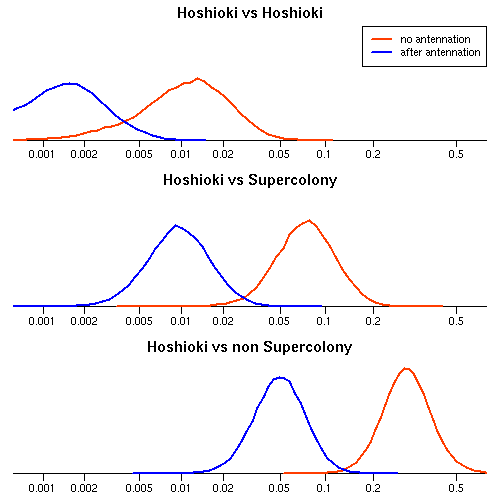
同巣の場合にくらべて,
相手が巣が異なる個体だと,
antennation されるリスクは 2.4 倍,
攻撃されるリスクは 14 倍上昇
-
これが石狩浜スーパーコロニー外から来たとすると,
上の値にさらに
antennation されるリスクは 1.2 倍,
攻撃されるリスクは 5.8 倍上昇
-
拉致られアリの接触回数 (1-5 回)
が増えるにつれ,
攻撃されるリスクは 1 回ごとに 1.2 倍ずつ増える
(5 回目だと 3 倍!)
……
これが上でいう「疲れ」効果
-
巣から 30 m 離れたところから拉致されてきたアリが
攻撃されるリスクは 2.3 倍上昇
-
逆に,
いったん antennation されると
攻撃されるリスクは 0.1 倍に
……
antennation されてる間に逃げ出すスキがあるから?
-
このデータでは
「特別にアヤしまれる・攻撃されるアリあるいは出身巣」
は見つけられなさそう
……
つまり異なる巣出身かどうか,
スーパーコロニー外から来たかどうか,
が判断の上で重要
-
antennation に関しては「実験場所」の効果はとくにないけど,
攻撃しやすさに関しては
星置の巣のワーカーを 1 とすると,
オッズ比が厚田 = 1.7, 忍路 = 0.8, 定山渓 = 27
となっていて定山渓のいよーな攻撃性が
……
と言いたいところで平均値 (中央値もだいたい同じ)
はこうなってるんだけど,
じつはこの「実験場所」効果の事後分布はどれも
95% 信頼区間内にゼロを含んでいる
(おそらく厚田・忍路・定山渓は観察回数が格段に少ないため)
ので,
そういうパラメーターであるという保留つき
-
いきなり「計算おわった」脱力感が.
いや,
このアリの行動実験解析にしても事後分布図を作ったり,
説明を書いたりとかいろいろあるんだけど
……
とりあえず A801 院生部屋 Dell 機はまたしばらく休眠させようか.
自分の机の上から
sudo shutdown -h now
発令してから,
主電源のスイッチを切りにいく.
-
気分転換のため,
八甲田「脱」割算値メイルかき.
これまたいろいろと考えてしまって,
なかなか時間がかかる.
-
いろいろ論文ダウンロード,
とか.
-
データ解析こんさるメイル書き.
一般化線形混合モデル計算関数
glmmML()
(あるいは R の
help)
でいろいろ出てくる「えらー」について.
こういう例で説明してみましょうか……
> library(glmmML)
> y <- rpois(100, exp(1 + rnorm(100, 0, 1)))
> id <- 1:100
> glmmML(y ~ 1, cluster = id, family = poisson)
Call: glmmML(formula = y ~ 1, family = poisson, cluster = id)
coef se(coef) z Pr(>|z|)
(Intercept) 1.07 0.116 9.18 0
Standard deviation in mixing distribution: 0.917
Std. Error: 0.0923
Residual deviance: 241 on 98 degrees of freedom AIC: 245
これは「切片」だけモデルで,fixed effects の部分が 1 (正しく推定されてます
ね),そして linear predictor にくっつく平均 = ゼロ かつ SD = 1 の正規分布が
random effects をあらわしています.この SD の推定値はご指摘のとおり Standard
deviation in mixing distribution: に 0.917 と (だいたい正しく) 推定されて示
されていますね.蛇足ながらその下の Std. Error: 0.0923 は上の 0.917 なる推定
値の推定のばらつきです.
上の例で,私のところで生成した乱数例で計算してみると,mean(y) = 4.4 の
var(y) = 24.8 となりポアソン分布からの overdispersion になっており,これを説
明するためには random effects が必要です.上の例では linear predictor にくっ
つく平均ゼロの正規分布を使ってそれをモデル化していて,この正規分布の SD を上
のように推定してます.
さてさて,この random effects が大きかろうがゼロに近かろうが,切片の推定値に
「自信のなさ」はつきまとうものであり,これはデータ数の大小その他あれこれでき
まります.これが推定値の se(coef) となります.
1945 研究室発.
2000 帰宅.
晩飯.
[今日の運動]
[今日の食卓]
- 朝 (0930):
米麦 0.7 合.
ネギ・豆腐の味噌汁.
キャベツ・ニンジン・キュウリのサラダ.
- 昼 (1400):
研究室お茶部屋.
イソップベイカリーのバゲット.
リンゴ.
- 晩 (2100):
マカロニ.
ピーマン・トマト・バジル・豆腐のソース.
キャベツ・ニンジン・キュウリのサラダ.
イソップベイカリーのバゲット.
2007 年 06 月 13 日 (水)
-
0600 起床.
よく眠れなかった.
コーヒー.
朝飯.
0800 自宅発.
晴.
予報では今日も暑くなるそーで.
0815 研究室着.
-
昨日のアリ計算結果とかみなおしてるうちに,
計算方式をびみょーに変えてやりなおしたほうがよいかも,
という気がしてきた.
今のモデルでは
Non.supercolony
なる説明変数があるんだけど,
これは拉致られアリの出身地が石狩浜スーパーコロニーの外
(つまり厚田・忍路・定山渓) かどうかをあらわしているのだが
……
-
うーむ,
解析の意図としては,
スーパーコロニー内外の対決かどうか,
をあらわす変数に変えたほうが良いのかも,
という気がしてきた.
ほとんどの実験 (330 回中 240 回) はスーパーコロニー内 (星置)
で行われているので
Non.supercolony
が「内外対決」あらわすわけだが,
残り 90 回はスーパーコロニー外 vs 外対決なんだよね.
-
懸念してるのは
Non.supercolony
ではなく
Sc.vs.nonsc
といった変数に変えた場合,
推定計算全体に悪影響およぼしがちな定山渓での実験結果の
統計モデリングがさらにハタ迷惑なものになりそう,
というあたりなんだけどね
……
-
とりあえず,
ものの試しとて再計算.
計算まち時間に岩倉さん修論をみなおす.
-
修正した階層ベイズモデルでの MCMC 計算,
やっぱり収束が悪くなった
……
そうか,
Sc.vs.nonsc == 1
だと実験場所は (石狩浜スーパコロニー内の)
星置に決まっているから
「複数パラメーターの見わけがつかない」
状態におちこんでしまうわけだ
……
で,
定山渓アリの突出した攻撃性にひっぱられて,
星置の攻撃性がふらふらしがちになる,
と.
えーい
……
-
「星置が実験場所である」
効果をゼロとおくか.
つまりここを基準にして他の実験場所の影響が決まる,
と.
名義変数の
levels 変換わざはこう,
と.
d$nest <- factor(d$nest, levels = c("hoshioki", "atsuta", "oshoro", "jozankei"))
-
いやー,
このやりかたはまずいかもしれないな.
「実験場所」効果があたかも fixed effects 的に
……
うう,
まあ,
fixed 的でもいいか.
ということでこのまま計算を強行.
for (nest in 1:N.nest) { ... }
としてたところを
for (nest in 2:N.nest) { ... }
として星置はゼロで固定,
と.
はてさて,
どうなることやら.
-
MCMC 計算,
キレイに収束してしまった
……
しかし,
やっぱりモデルが恣意的なかんじになるので,
これはやめとこう.
-
で,
fixed effects のふらつく範囲を制約するような
hyper parameter を導入してみる,
と
……
いや,
これもあまり意味ないとわかった.
-
いろいろ苦闘してるとすでに 1400 すぎ.
MCMC 計算を走らせといて,
昼飯調達の旅にでる.
あいかわらず天気よい.
計算待ち時間に読んだ岩倉さん修論の内容をアタマの中で反芻しつつ
モデリングを検討していると,
うまくいきそうな策をひとつ思いついた.
とりあえず昼飯買って,
研究室にもどって昼飯.
-
さて,
モデリング改善策だが
……
けっきょく,
端的に言えば,
データの中では多数派である海ぞいのアリたちは
皆のんびり暮らしているのに,
少数派である山ぶかい定山渓の孤独の中で
アタマがヘンになって凶暴化して噛みつきまくるアリどもが
統計モデリングを困難にしているのである.
今までこの凶暴性をなんとか random effects に押しこもうとしたけれど,
その試みはことごとくしくじりつつあるところだ.
-
では,
どうすれば良いかといえば,
この凶暴性を fixed effects として説明すればよい,
というあたりまえのコトに今さらながら気づいた.
定山渓以外のコロニーは polydomous である
(ワーカーが巣間をいききする)
のに対して,
定山渓ひきこもりアリどもはワーカーの巣間移動ナシ,
と岩倉さん修論に書かれている
(ひきこもりだけど多女王 (polygynous)?).
-
つまり
……
えーと polydomy でないのは
monodomy
でよいのかな
……
拉致られアリ出身地を特徴づける Monodomy
なる説明変数を追加してやればいいだろう.
完全に定山渓ねらいうちのイヤなモデリングだけど,
ともかくこの凶暴アリどもにふりまわされるのは,
もううんざり,
ということで.
-
このモデリングはうまくいった
……
しかし fixed effect parameter を増やしてしまったことで,
攻撃確率に関係するパラメーターたちの収束が悪くなる.
-
このあたり不自然にならぬよう,
超事前分布をひかえめにしたり,
可能なかぎり fixed effect parameter 数を減らしたりする工夫を
……
計算まち時間にうろうろしてると,
永光さんがお茶部屋におられたので,
アリその他雑談.
おかげさまで,
アタマが少しばかり整理される.
-
まあ,
こんなものでよろしかろー,
という結果ようやく得られる
(事後分布表).
とうぜんながら,
というか結果の解釈は昨日とほとんど同じ.
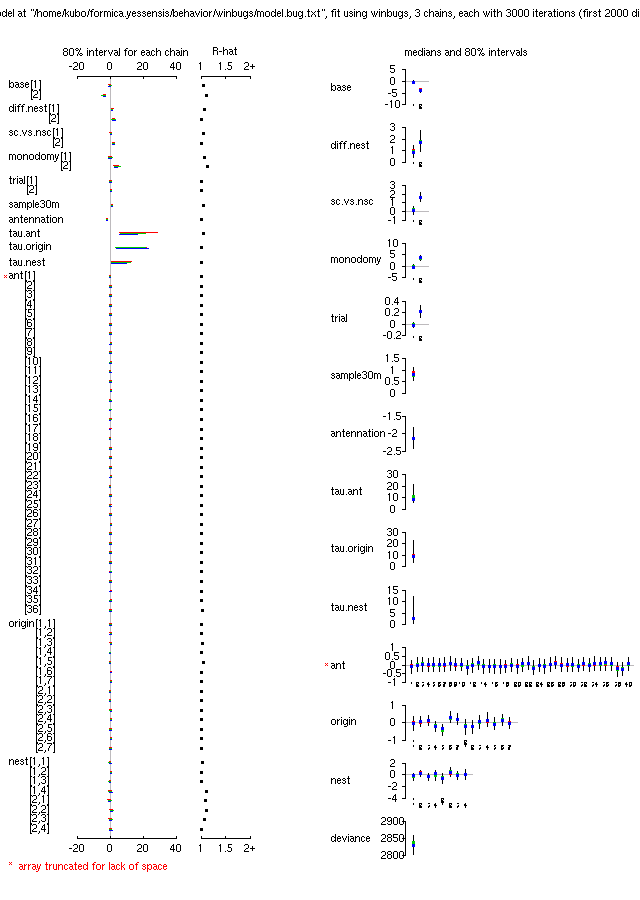
-
とくに何かパターンあるわけでもないけど,
fixed effect parameter 事後分布
pairs() 図.
時刻はすでに 1820.
今日はこの計算と,
岩倉さん修論読みとこんさるメイル書き,
で終わりつつあるな.
-
1940 研究室発.
2000 帰宅.
晩飯の準備.
晩飯.
-
[今日の運動]
-
[今日の食卓]
- 朝 (0700):
米麦 0.6 合.
ネギ・ニンジン・卵の炒めもの.
- 昼 (1420):
研究室お茶部屋.
米麦 0.6 合.
春雨スープ.
- 晩 (2130):
米麦 0.7 合.
ジャガイモ・ニンジン・タマネギ・グリンピース・挽肉の煮物.
プチトマト.
コマツナ.
2007 年 06 月 14 日 (木)
-
0820 起床.
コーヒー.
朝飯.
0940 自宅発.
曇.
今日から少しすずしくなるはず.
1000 研究室着.
-
午前中はアリ計算まわりの整理,
アリ論文ダウンロードなんかで終わってしまった.
-
昼飯.
今日は院生密度がすごく低いので,
お茶部屋で甲山さんとのんびり雑談.
-
データ解析こんさるメイルかき.
-
昨日の夕方得られた計算結果,
(プログラムその他はちゃんと backup してたのに)
まぬけなことに
save(ant.bugs, file = "ant.bugs.RData")
しとくの忘れてた!
われながらまぬけだ
……
1000 秒かけて再計算.
脱力.
-
……
と思ったら,
ant.bugs.RData
ではなく
ant.bugs
というファイル名でちゃーんと保存していた,
と発見.
さらに脱力.
いやはや.
-
てなかんじであまり進捗せず,
1815 研究室発.
ちょっと雨.
1830 帰宅.
洗濯.
晩飯.
-
……
あ,
いきなり気絶するみたいに寝てしまった.
真夜中すぎに目をさます.
-
[今日の運動]
-
[今日の食卓]
- 朝 (0900):
米麦 0.6 合.
ジャガイモ・ニンジン・タマネギ・グリンピース・挽肉の煮物.
コマツナ.
- 昼 (1220):
研究室お茶部屋.
米麦 0.8 合.
ジャガイモ・ニンジン・タマネギ・グリンピース・挽肉の煮物.
コマツナ.
- 晩 (2010):
米麦 0.7 合.
ジャガイモ・ニンジン・タマネギ・グリンピース・挽肉の煮物.
コマツナ.
2007 年 06 月 15 日 (金)
-
0630 起床.
コーヒー.
朝飯.
0740 自宅発.
雨.
0755 研究室着.
-
セミナーの予習.
-
1030 より
研究室セミナー,
今日は田辺さんで樹木
(といっても seedling だの sapling といった小さな個体)
の地上部-地下部の重量分配やカタチを調べました,
そうするとこういうハナシになります,
といった論文の紹介.
- Guerrero-Campo J., Fitter A.H. (2001) Relationship between root
characteristics and seed size in two contrasting floras. Acta
Oecologica 22: 77-85
(abstract) Plants vary greatly in root system characterisitcs,
but the causes of this variation are poorly understood. We hypothesized that root
system size is closely linked to the plant's ecological strategy,
and that seed size is correlated with root diameter, as a result of anatomical
constraints. We analyzed the relationships between root characteristics
- root depth, root diameter and root type - and other plant attributs
in more than 300 plant species from two ecologically and geographically contrasted
areas: Britain and NE Spain. We used statistical tests that
took into account phylogienetic patterns in the data.
Apart from plant life span, only plant height and seed size were related to root size
in the adult plants. Plant species with shallow or thin main roots had smaller seeds
than species with deep or thick main roots, and species with taproots
was related to plant height, but this association was weaker than that between
seed size and root depth. Root depth explained a siginificant
proportion of the variation in seed weight, independently from life form or dispersal
mode and, in some cases, more than either of them. These
results suggest that traditional ecological explanations do not adequately explain the
relationship between seed size and plant adult size, and that
there will be other complementary explanations. In particular, we propose that the
relationship between seed size and plant height is secondary.
The putative causal sequence is that deep-rooted plants (which are generally taller)
have large seeds because of allometric and developmental
constraints that mean that only large seeds can produce the thick roots that
can grow rapidly to depth.
- Kohyama T., Grubb P. (1994) Below- and above-ground allometries of
shade-tolerant seedlings in a Japanese warm-temperate rain forest.
Functional Ecology 8: 229-236
(abstract) ... 4. Interspecific differentiation in below- and above-ground dry mass is not
correlated with that in above-ground allometries at the seedling stage or with tath in
above-ground allometries at the sapling stage with developed branching architecture.
Allometric `design' in architecture at the sapling stage (related to earlier or later
canopy-gap formation) is variously combined with that at the seedling stage (related
to the microsite of establishment).
- Nishimura T., Suzuki E. (2001) Allometric differentiation among
tropical tree seedlings in heath and peat-swamp forests. Journal of
Tropical Ecology 17: 667-681
(abstract) ... We concluded that seasonal water limitation brought about the
convergence in seedling morphology within heath forest because average understory
irradiacne and soil nutrient concentration were assumed to be similarly low both
forests.
- Paz H. (2003) Root/Shoot Allocation and Root Architecture in
Seedlings: Variation among Forest Sites, Microhabitats, and Ecological
Groups. Biotropica 35(3): 318-332
(abstract) I analyzed patterns variation in root mass allocation and root morphology among
seedlings of woody species in
relation to environmental factors in four Neotropical forests.
Among forests, I explored the response of root traits to
sites varying in water or nutrient availability. Within each forerest,
I explored tha plastic response of species to defferent
microhabitats: gaps and understory. Additionally, I explored evidence for life history
correlation of root and shoot trailts by comparing species differring in their
successional group (light-demanding [22 spp.] or shade torelant [27 spp.])
and germination type (species with photosynthetic cotyledons or species
with reserve cotyledons). At each forest site,
young seedlings from 10 to 20 species were exavated.
A total of 55 species was collected in understory conditions
and 31 of them were also collected in gaps.
From each seedloing, six morphological ratios were determined.
...
Overall, these results suggest that among Neotropical species,
root allocation and root morphology of seedlings reflect plant adjustments to water
or nutrient availability at geographic and microhabitat scales.
In addition, life history specialization to light environments
is suggested by differences among groups of species in their allcation to roots
and in their root morphology.
- Yamada T. et. al (2005) Tap-root depth of trpical seedlings in
relation to species-specific edaphic preferences. Journal of Tropical
Ecology 21: 155-160
(abstract)
...
This trait was fixed independently of the edaphic conditions win whiich the trees
actually grew. Finer-textured soils generally have better water-holding capacity;
we concluded that seasonal water limination would be more severe
in sandy soils than in clay-rich soils, and that water limination would proveke
convergence in seedling root depth
amongh species with the same edaphic preferences.
まあ,
基本的に,
静的な記述で成長過程は調べないんだよね
……
(その場における甲山さんのご指摘なんだけど)
落葉・細根の消失といった「呼吸」のデータはなかなかないからね.
-
まあ,
このあたりの解析すすめるためには,
またいろいろと勉強せねば
……
-
北大構内昼飯調達の旅.
雨はいつのまにかあがっている.
研究室にもどって昼飯.
-
ひさびさに生態学会さーばーネット雑用.
ちょっとアカウント & ディレクトリ作るだけ,
だったんだけど.
-
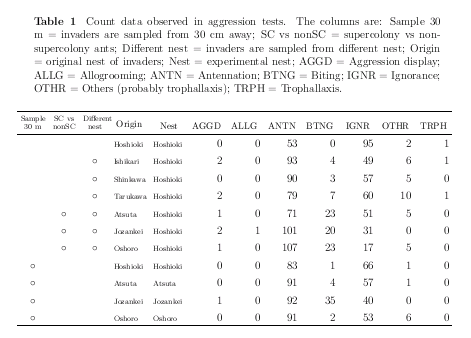
拉致られアリ実験の階層ベイズ推定結果の Table
作り
……
じつは昨日からひっそりと苦闘してる.
うーん,
こんなもんかしらん?
いつものごとく R
& LaTeX コンビネイション自動作表わざ.
-
カウントデータの Table はレイアウトに苦闘する.
-
2010 研究室発.
2035 帰宅.
体重 67.6kg.
晩飯.
-
明日はなぜかまた甲山さんと
定山渓天狗岳
にのぼりにいくことに
……
-
[今日の運動]
-
[今日の食卓]
- 朝 (0720):
米麦 0.6 合.
コマツナ・マイタケ・卵の炒めもの.
- 昼 (1310):
研究室お茶部屋.
バターロール.
リンゴ.
- 晩 (2200):
米麦 0.8 合.
ネギ・ショウガ・豆腐の炒めもの.
コマツナ・マイタケ・卵の炒めもの.
2007 年 06 月 16 日 (土)
-
0630 起床.
よくねむれなかった.
眠い.
ウスいコーヒー.
朝飯.
-
0725 自宅発.
快晴.
甲山さんが車で当家ちかくまで来てくださったので,
それに乗せてもらって
定山渓天狗岳
にむけて出発.

[08:33 道道 95 からみた定山渓天狗山]
山頂岩峰部のせいで独特の山容になっている.
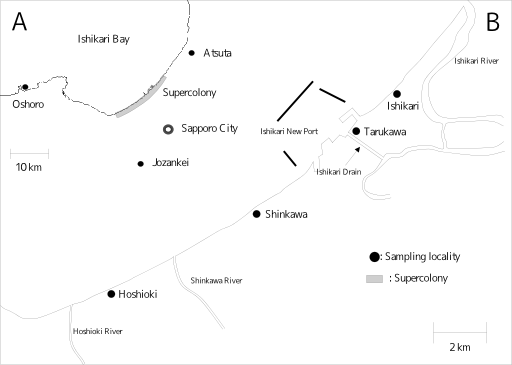
周辺の位置関係はこの
地図
参照.
0835 白井川二股の橋をわたった駐車場着.
すでに先行パーティー多数.

[09:00 白井川ぞい林道を下る]
0850 駐車場発.
駐車場からのびる林道はゲイトで閉鎖されてたわけだが
……

[09:18 天狗岳登山口]
登山口ににはなぜか自家用車が.
どうも下流がわの橋から入ればここまで進出できるらしい.
ここから左側に登山道つづき,
しばらく熊ノ沢左岸をのぼっていくことに.
これは 1/25000 地図とは異なる
(地図がまちがっている).

[09:32 最初の渡渉点]
あとで問題となる「滝」までは比較的ゆるやかな登りがつづく.

[09:47 まちがい尾根にふみこむ]
次の次の写真にある滝を (下流からみて) 左のほうの尾根に
登って高まこうとしたわけだが
……
これは大失敗で,
こっちに来てはいけない.
かなりの急登つづく.

[09:59 かなり見晴らしよいところまで]
登ってしまって,
「なんとか突破できないか」
とササ原を迂回したりみょーなアカエゾ植林ベルトをうろうろしてから,
「やっぱりここはダメだ」とあきらめて撤退
(あとで山頂からこのササ原の広漠ぶりがわかり,
これの突破計画は無謀そのものだったと反省).
この撤退,
route finding にじつに「気あい」を必要とするもので
……

[10:47 問題の滝にもどれた]
まったく奇跡的にもとの熊ノ沢ぞいのルートに下降できた
……
もどれた瞬間,
がっくりと脱力.
道をまちがえると体力・気力を使いきるよ.
しばらく上流にあるくと問題の滝に再開
(つまりまちがえて登った尾根とは別の尾根を下ったのである!).
ともかく,
この滝は下流からみて右側の岩面をトラヴァースぎみに登るのが正解.
嗚呼.

[10:57 次の滝]
問題の滝からちょっと登ると,
ゴルジュというほどでもないけど,
沢は岩壁にはさまれていて雪などが詰まっていたりする
(もうちょっと上流では滝になっている).
ここは沢ぞいではなく高まくのが正しいルートで,
下流からみて左側を登っていく.

[11:28 標高差 400 m の急登]
で,
その滝の横を突破すると熊ノ沢の水量はいよいよすくなくなり,
沢の源頭部というべき地帯をひたすら登っていくことになる.
浅い水流を何度も渡渉したり.
山頂までの標高差はおよそ 400 m.
ひたすらに急登である.

[11:58 天狗岳 I 峰西面の岩壁]
沢の源頭部急登をひたすら登っているとこの岩壁がいきなり立ちふさがる.
もちろんこんなところは登れないので,
岩壁の下 (つまり I 峰基部) を時計まわりにまいていく.

[12:14 fixed rope つきルンゼ]
I 峰の北にまわりこむと I-II 峰間のコルにつく.
そこから I 峰にむけて最後の急登になる.
途中に 20 m ほどのルンゼがあって fixed rope が残置されてる.
ロープは腐れぎみだし,
傾斜もそれほどきつくないので岩場に慣れたヒトなら
ロープとかに触らないほうがさっさと登りおりできる.
ここを登ると I 峰西側ジャンダルムと本峰のコルみたいなところに出る.
1215 せまい山頂着.
めでたしめでたし.

[12:27 山頂から南東方向をみる]
ダム湖よこ手前のピークは小天狗岳.
その向こうは
昨年 10 月
に行った札幌岳とか
(札幌岳 - 空沼岳 - 漁岳 と並んでる).

[12:28 山頂から南西方向をみると]
手前に無意根山,
そのむこうにカルデラ形の羊蹄山やニセコ連峰など.
ところで,
今日は風がよわかったせいか,
とくに山頂付近は虫がたくさん飛び回っていて,
なんともうっとうしかった.
甲山さんに虫よけスプレーおかりしたんだけど,
あまり効かない.
あ,
それからこういう日は日焼けどめが必要だな
……

[12:38 さっぽろ湖]
東面はこの巨大なダム湖が.
烏帽子岳・神威岳がダム湖のむこうに.
さらにその向こうには定山渓・八剣山が.
そう,
あの凶暴アリどもがサンプルされたという
……

[12:38 北東みると]
大学からもよくみえる「電波塔山」である手稲山が「ウラ」から見えておもしろかった.
このへぼい写真では何もわからんけど.
1250 下山開始.
急登ルートをひたすら下る.
昨日の朝まで雨ふってたはずなんだけど,
きれいさっぱり乾いていて助かった
……
泥濘の急な山道くだりはけっこうしんどいんで.

[15:16 駐車場に無事にかえりつく]
復路は往路のような大冒険もなく,
無事にかえりついた.
おつかれさまでした.
1530 駐車場発.
1543 定山渓より札幌市内よりの小金湯温泉へ.

風呂からあがったところ.
体重 67.0 kg.
今日は水を 1.2 リットルぐらい飲んでしまったんだけど,
やはり発汗量はそれをはるかに上まわっていたんで,
いつもよりやや低めの体重測定値となった.
-
また甲山さんに自宅ちかくまで送っていただいた
(ありがとうございます).
1710 無事に帰宅.
洗濯.
ばてる.
-
晩飯どうしようかと思ったんだけど
……
買いものにはいかずに冷蔵庫の残りものですます.
-
[今日の運動]
-
[今日の食卓]
- 朝 (0645):
米麦 0.6 合.
ネギ・ショウガ・豆腐の炒めもの.
- 昼 (1220):
定山渓天狗岳山頂.
にぎり飯 2 個.
- 晩 (1940):
米麦 0.7 合.
ネギ・卵炒飯.
ニンジン・タマネギ・高野豆腐のシチュー.
2007 年 06 月 17 日 (日)
-
0730 起床.
よく眠れたのかそうでないのか
……
コーヒー.
朝飯.
-
当家の劣悪な小型冷蔵庫 (一室式)
内の掃除.
こういう一室式の欠点のひとつは上部に製氷室なるものがあって,
ここにすごく着氷 & 解氷して庫内がべとべとになったりするする,
ってあたりにあるわけで.
朝飯準備中から電源とめて,
氷をはがす.
-
で,
掃除なんかしてると周辺の床もけっこうぬれてしまったので
……
ひさびさに床掃除を決行.
けっきょくあちこち掃除して午前中はこれで終了.
うう.
-
ばてたので手ぬきな昼飯.
怠業.
-
1510 自宅発.
晴.
1525 研究室着.
-
ふーん,
各務原にアルゼンチンアリ
……
-
アルゼンチンアリではないけど,
同じく多女王・多巣性のエゾアカヤマアリの作図プログラミングのつづき.
R + LaTeX の
picture 環境わざ,
と.
-
さて,
この拉致られアリいじめ攻撃の階層ベイズモデルを MCMC 計算して
得られる事後分布図のほうだけど
……
うーむ,
いわゆる fixed effects のほうは
一昨日にすでに Table にしちゃってるわけで
……
-
とすると,
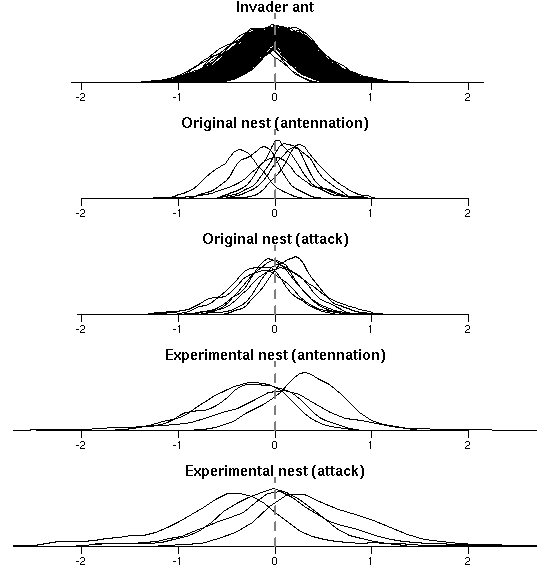
Table 化むずかしい random effects の事後分布図でも作ればいいのか?
つまり拉致られアリ個体,拉致した巣,拉致られ先の巣の効果,
ということになる.
巣に関しては
「しつこい触角チェック (antennation)」
「噛みつきとか攻撃行動」
それぞれに推定されてるんだよねえ.
-
その「下描き図」みたいなものを作ってみる.
ふーむ,
突出してヘンな個体・巣はない,
ということかしらん?
定山渓の凶暴アリどももいまや
Monodomy
なる fixed effects で「説明」されちゃってることになるわけで
……
-
空腹になってきたので本日はここまで.
1850 研究室発.
1905 帰宅.
晩飯の準備.
晩飯.
-
[今日の運動]
-
[今日の食卓]
- 朝 (0830):
米麦 0.7 合.
ピーマン・ネギ・卵の炒めもの.
ニンジン・タマネギ・高野豆腐のシチュー.
- 昼 (1220):
米麦 0.7 合.
レトルトパウチドカレー.
キュウリ.
- 晩 (2040):
米麦 0.7 合.
紫アスパラガス・タマネギ・ネギ・ピーマン・ニンジン・
ショウガ・ニンニク・イカの炒めもの.
キャベツ・ニンジン・タマネギ・キュウリ・トマトのサラダ.
2007 年 06 月 18 日 (月)
-
0730 起床.
コーヒー.
朝飯.
朝からアリ作図修正にはまりこんでしまった.
0945 自宅発.
曇.
1000 研究室着.
-
昨日つくりかけてた
random effects なパラメーターの事後分布図,
清書版.
R では 5 個の「部品」だけを
postscript(..., file = "special")
で EPS 出力し,
あとは LaTeX 内の picture 環境内外でレイアウトする.
-
この図からわかることは,
アリ拉致 & ケンカ実験では個体差・出身巣・実験巣それぞれの
「効果の正負」はことごとくよくわからんけれど
(まあこの大統領方式の実験計画からすると妥当だと思う),
しかしながらその推定の uncertainty は無視できませんよ
(「なかったこと」にはできない),
というものだ.
-
fixed effects の事後分布図かこーかどーか,
という判断がよくわからなくなったので,
気分転換にアリ拉致 & ケンカ実験図をつくってみる.
おびえたキモチで逃げまわる拉致られアリを
ちくちくと攻撃する矢印を描いていると,
なぜかしらココロの平和が実現する.
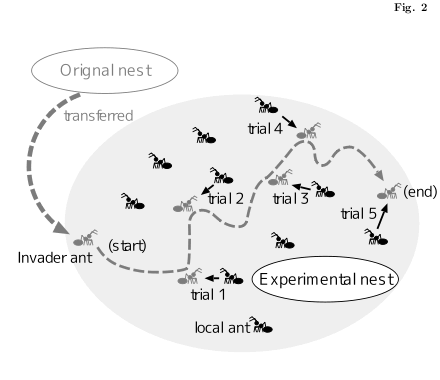
-
じゃあ,
ついでに岩倉さん作成の地図も,
と変換にとりくんでみたんだけど
……
くそう,
MS おひすファイルから OpenOffice.org (ver 2.0.4)
を使って図を取りだせるんだけど,
こいつを LaTeX で読めるカタチで保存できん
……
-
困ったので院生の PC の poorpoint 借用すれば何かうまい
export ができるか,
と試みてみたんだけど,
WMF ってのはぜんぜん他のソフトウェアでは読めんね.
-
また PC れんたる料として計算したうけを約束させられてしまったので,
いそいで昼飯をくう.
-
で,
昼飯後に宮田さんの苫小牧広葉樹の当年生シュート重量分配の階層ベイズモデル.
じつはこのへんの樹木の葉っぱってけっこう虫にくわれてるんだけど,
今までのデータ解析ではこれを無視してた.
つまり「葉重量」といっても虫に食われたあとの重量だったわけで
……
-
で,
宮田さんが葉っぱスキャナ画像みなおして,
「食害度」 0-4 なるものをシュートごとに測定しなおして,
それをつかって Bayesian なモデリングしなさい,
という問題.
これはどうすればよいかというと,
たとえばあるシュートが「食害度 1」なら,
そのシュートの「葉が食われた割合」の事前分布を
範囲 0.01 - 0.25 の一様分布とおいて
(
dunif(0.01, 0.025) てなかんじで),
そこから事後分布だしなさい,
とするわけ.
これを 13 樹種 500 シュートについていちいち計算していく,
と.
-
で,
以前の計算プログラム改良,
食害の効果をいれて MCMC 計算やりなおす
……
収束が速くなり,
「測定誤差」は格段に小さくなった.
というのも今まで虫くいとかで葉重量おかしな値になってたなどなどの
矛盾のすべて
をこの
「測定誤差」とやらに押しつけていたためである.
-
ただし,
多くのシュートで「まあ虫くわれ割合は大きくてもよし小さくてもよし」
みたいに事前分布の範囲内でうろうろしてくれるせいなのか,
定数係数パラメーターの収束がわるい.
-
そこで (他のパラメーターの収束はよいことから)
この定数係数パラメーターだけが「ゆっくりうねる」状態にあるのだろう,
ということで総 MCMC 計算 step 数は 4000 に固定したまま,
サンプリングは最後の 3000 MCMC step 15 とばしの
200 個のトリプルチェイン,
でやってみる.
Dell デスクトップ機で 630 秒ぐらい.
これでうまくいった.
以前は 60000 MCMC step も必要としてたのに比べれば
(半年ほど前に苦闘してました),
格段に改良された.
-
「虫にどれだけ食われたか」の事後分布はシュートによってけっこう異なる.
同じ「食害度 1」であっても 1-25% の範囲いっぱい使って
ひたすらふらふらするのもあれば,
ぜったいに 20% 以下には落ちない,というようなのもまじっていて,ですね
……
なかなか興味ぶかい.
-
あと,
「樹高がたかくなるほど虫に食われてしまう」
という樹種がいくつかあったんだけど,
今回の食害くみこみモデリングでこういう偏りもたらす要因をうまく除去できた,
と.
めでたしめでたし,
すばやく一件落着.
とはいえ,
時刻はもうすぐ 1600.
-
エゾアカヤマアリ作業に復帰.
岩倉さんが気合いいれて作った調査地周辺の地図を
MS おひすという牢獄からサルヴェイジする作業のつづき.
けっきょく
-
Linux 上の OpenOffice.org で開く
-
「印刷」で postscript file 出力
-
お茶部屋 iMac にもっていって,
preview で PDF に変換
-
iMac 上の Illustrator CS1 で開く
-
なぜか地図の一部がこわれてるので,
PDF 変換前の postscript file を Illustrator
で開いてこわれてる部分をこぴぺ修復
-
Illustrator CS から EPS 出力
-
Linux 上の LaTeX file 内の
\includegraphics
でとりこむ
-
make pdf
とゆー,
わけのわからぬ呪術的コンボでどーにかこーにか
……
-
まあ,
あとで時間あったら Tgif とかで trace して
作りなおしたほうがよさそうだな
……
-
岩倉さん修論をみなおす
……
さてさてここまでの階層ベイズモデルによる解析をみなさんに
どう説明したものやら.
-
と考えてると,
よその研究室の院生のデータ解析こんさる.
dhyper()
すなわち超幾何分布 (hyper geometric distribution)
つかう問題だった.
空腹になってきたし,
帰るとしますか.
1950 研究室発.
2005 帰宅.
体重 67.8kg.
晩飯.
-
[今日の運動]
-
[今日の食卓]
- 朝 (0800):
バターロール.
キャベツ・ニンジン・タマネギ・キュウリ・トマトのサラダ.
- 昼 (1330):
研究室お茶部屋.
米麦 0.6 合.
レトルトパウチドカレー.
- 晩 (2120):
米麦 0.7 合.
チンゲンサイ・タマネギ・卵の炒めもの.
コマツナのゴマあえ.
2007 年 06 月 19 日 (火)
-
0740 起床.
コーヒー.
朝飯.
0910 自宅発.
晴.
0925 研究室着.
-
アリ作図のつづき.
こういう図をつくってみる.
電気鼠もちいぬ LaTeX 作図をやってみたわけだが
……
こういう
LaTeX コード
になってしまい,
こうなるとわかっていたら,
もっと「長さ変数」とか設定しまくるプログラム然としたモノ
にすればよかったかも,
と.
あるいはこういうコードを生成する R プログラム作ればよかったかな.
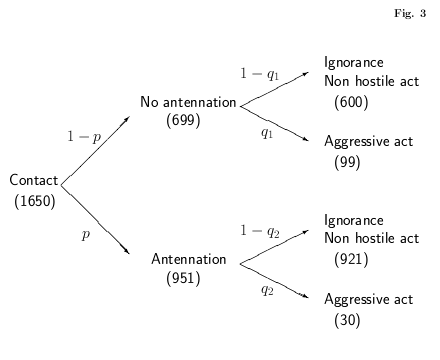
-
じつはこの図で表現されているモデルには問題があって
……
「ホントに antennation が先か?」
ということなんだよね.
岩倉さんに確認しておかなくては.
まあ,
おそらく攻撃に関しては攻撃→その後 antennation ってのは
ほとんどないだろう,
と思うんだけど.
-
1030 より
研究室セミナー,
今日は木村君で青森県
仏沼
の火入れ処理に対するヨシその他植物の応答しらべます,
という自分の研究計画の紹介.
湿原のヒトたちは,
地面の PH とか測るの好きだねえ
……
しかし
(空間的にそれほど離れてない場所で測定された)
こういった測定値が現象の説明に有用だった,
という研究例はあるのだろうか?
-
やっぱり何か周辺 (marginal) 事後分布図だしたほうがいいよなー
……
と検討.
-
しかしよくわからぬまま昼飯.
-
ゐんばぐす出力をよみこんで,
R の中で条件つき事後確率を計算するプログラム作り.
330 (個体) × 600 (MCMC step)
の行列を 3 つ
(antennation 確率,antennation ナシの場合の攻撃確率,
antennation 後の攻撃確率)
を生成させる.
-
1500 より 70 分ほど動物生態の藤原君とエゾアカヤマアリ相談.
統計モデルについて説明してみると,
用語でまずい点に気づいたり,
あるいはちょっとヘンかもしれない点を指摘してもらえた.
-
「antennation が先」
というモデルはどうやら正しいみたい
-
nest / colony はちゃんと区別して使わんといかん
……
しかし polydomy の場合はあまり厳密にできんよーな
-
「このコロニーは supercolony の一部」
ってのは,
じつはケンカする・しないで supercolony
かどうか定義されてるんだよね
……
まあ,
「supercolony ということにされてたコロニー」
とでもしますか
-
supercolony を隠れ変数であつかう,
ってのもおもしろいアイデアかもねー
……
といったことども留意しつつ,
さっさと計算メモまとめたほうがよさそう.
-
で,
昼飯前後から苦闘してる事後確率分布図,
だいたい部品 (自作
function())
がそろってきたので,
ちょっと作図してみる.
-
横軸は対数軸ならぬ「logit 軸」.
ろじっとについては
粕谷さん馬車馬ぶろぐの
エントリー
参照のこと.
-
1925 研究室発.
1940 帰宅.
晩飯の準備.
晩飯.
-
おうぷんおひすで
EPS export
できるとのご教示で,
これって ver 2.2.* 系なんだろな
(2.0.* 系では PDF export しかできない),
download & install めんどくさいな,
と思いつつもとりあえず
sudo apt-get remove openoffce.org
とアンインストール命じて
(Vine Linux 版における最新版 RPM package は
2.0.4 ではなく 2.0.3 だった!),
ring server
2.2.1 をダウンロード.
これは汎用 RPM になってて,
必要なコンポーネントだけ sudo rpm -Uvh
できる.
-
たしかにおうぷんおひす 2.2.1 だと,
PDF だけでなく EPS その他で export できますなあ
……
おお,
SVG
export もできるってことは
inkscape で import できるわけで
……
これはまあまあうまく export-import できてるように見える.
まあ,
修正が必要なら inkscape でできそう,
ということで.
それなりに手作業による補正が必要そうだけど.
そうか,
SVG なら Perl とか R とかで変換できるかも!
-
[今日の運動]
-
[今日の食卓]
- 朝 (0810):
米麦 0.7 合.
ネギ・豆腐の味噌汁.
トマト.
コマツナのゴマあえ.
- 昼 (1250):
研究室お茶部屋.
食パン.
リンゴ.
- 晩 (2120):
米麦 0.7 合.
キャベツ・レタス・モヤシ・ニラ・ニンジン・ショウガ・
ニンニク・ブナシメジ・豚肉の炒めもの.
コマツナのゴマあえ.
2007 年 06 月 20 日 (水)
-
0730 起床.
コーヒー.
朝飯.
0845 自宅発.
曇.
0900 研究室着.
-
アリ作図,
これで終わりか?
な事後確率密度分布図
……
まあ,
いろいろと discussion のネタは供給してくれてますね.
-
今までつくった図表の整理.
-
昼飯.
Nature 447, 791-797 (14 June 2007)
に
Nature's guide for mentors
なる「学術師匠指南」みたいな feature 記事が掲載されてる.
なかなかおもしろい.
-
事務からまた奇妙な通知が.
夏季休暇の計画的使用について(通知)
夏季休暇は,特別休暇として,7月から9月までの期間内における休日を除く連続
する3日間取得できることになっております。
夏季休暇の取得にあたっては,「職員が夏季における盆等の諸行事,心身の健康の
維持及び増進又は家庭生活の充実のため勤務しないことが相当であると認められる場
合」の趣旨から,夏季休暇の前後に年次有給休暇を使用し,可能な限り連続した休暇
を取得されるようお願いします。
なお,夏季休暇を申請する場合は,「特別休暇簿」を提出していただきますが,
「理由欄」には,「夏季休暇」と記入願います。
明日の統計こんさるにそなえて勉強.
そして食品中の細菌数増殖は
ちょっと奇妙な (現象論的な?) 集団成長モデルが使われてるよーで
……
先日,
日焼けどめもつけずに定山渓天狗岳にのぼったので,
今ごろ顔の皮膚がひりひりぼろぼろしてきた.
夏至もまじかの快晴の日に,
われながらなんともうかつなことよ.
拉致られアリちくちく攻撃統計モデリングの計算メモ書きのため,
また岩倉さん修論の深淵 (でぃーぷ) なる世界にはまりこんでみる
……
統計モデリングについて説明するにしても,
やったことを全部くわしく書いてもアリ研究室のヒトたちは
あまり読んでくれそうにないので,
得られた推定結果の一部を強調できるようなモデルの説明を考えている.
で,
岩倉さん修論よんでるとそのあたり整理されてくるわけだが
……
とりあえず,
定山渓の凶暴アリたちの
「antennation やったそのあとにさらに噛みつく」
という非道さについては世に知らしめねばならぬ,
ということでそういうことを強調する図を生成するよう
R 作図プログラムを改造する.
-
ばててきたので
2005 研究室発.
2020 帰宅.
体重 67.2kg.
晩飯の準備.
晩飯.
-
へー
……
押井守監督で
スカイ・クロラ.
なンか意外な組みあわせというか
……
押井監督で「飛ぶもの」といえば
ティルトローター
だの
オーニソプター
だのあるいはあの一群の奇妙なヘリコプター,
なかんずく
二重反転ローター
とか
交叉反転式ローター
みたいな
(つまり
フォッケ-アハゲリス
な) ふうがわりなモノばっかり
(あと黄色い飛行船ね),
という印象だったんだが
……
はてさて,
どうなることやら.
-
[今日の運動]
-
腹筋運動 30 ×
3 回.
腕立ふせ 10 ×
3 回.
スクワット 100 回.
-
[今日の食卓]
- 朝 (0810):
米麦 0.7 合.
キャベツ・レタス・モヤシ・ニラ・ニンジン・ショウガ・
ニンニク・ブナシメジ・豚肉の炒めもの.
- 昼 (1320):
研究室お茶部屋.
食パン.
リンゴ.
- 晩 (2140):
米麦 0.7 合.
コマツナのゴマあえ.
ネギ・ニラ・ニンジン・モヤシ・豆腐・煮干の味噌汁.
トマト.


{kind=link}