ぎょーむ日誌 2007-06-15
2007 年 06 月 15 日 (金)
-
0630 起床.
コーヒー.
朝飯.
0740 自宅発.
雨.
0755 研究室着.
-
セミナーの予習.
-
1030 より
研究室セミナー,
今日は田辺さんで樹木
(といっても seedling だの sapling といった小さな個体)
の地上部-地下部の重量分配やカタチを調べました,
そうするとこういうハナシになります,
といった論文の紹介.
- Guerrero-Campo J., Fitter A.H. (2001) Relationship between root
characteristics and seed size in two contrasting floras. Acta
Oecologica 22: 77-85
(abstract) Plants vary greatly in root system characterisitcs,
but the causes of this variation are poorly understood. We hypothesized that root
system size is closely linked to the plant's ecological strategy,
and that seed size is correlated with root diameter, as a result of anatomical
constraints. We analyzed the relationships between root characteristics
- root depth, root diameter and root type - and other plant attributs
in more than 300 plant species from two ecologically and geographically contrasted
areas: Britain and NE Spain. We used statistical tests that
took into account phylogienetic patterns in the data.
Apart from plant life span, only plant height and seed size were related to root size
in the adult plants. Plant species with shallow or thin main roots had smaller seeds
than species with deep or thick main roots, and species with taproots
was related to plant height, but this association was weaker than that between
seed size and root depth. Root depth explained a siginificant
proportion of the variation in seed weight, independently from life form or dispersal
mode and, in some cases, more than either of them. These
results suggest that traditional ecological explanations do not adequately explain the
relationship between seed size and plant adult size, and that
there will be other complementary explanations. In particular, we propose that the
relationship between seed size and plant height is secondary.
The putative causal sequence is that deep-rooted plants (which are generally taller)
have large seeds because of allometric and developmental
constraints that mean that only large seeds can produce the thick roots that
can grow rapidly to depth.
- Kohyama T., Grubb P. (1994) Below- and above-ground allometries of
shade-tolerant seedlings in a Japanese warm-temperate rain forest.
Functional Ecology 8: 229-236
(abstract) ... 4. Interspecific differentiation in below- and above-ground dry mass is not
correlated with that in above-ground allometries at the seedling stage or with tath in
above-ground allometries at the sapling stage with developed branching architecture.
Allometric `design' in architecture at the sapling stage (related to earlier or later
canopy-gap formation) is variously combined with that at the seedling stage (related
to the microsite of establishment).
- Nishimura T., Suzuki E. (2001) Allometric differentiation among
tropical tree seedlings in heath and peat-swamp forests. Journal of
Tropical Ecology 17: 667-681
(abstract) ... We concluded that seasonal water limitation brought about the
convergence in seedling morphology within heath forest because average understory
irradiacne and soil nutrient concentration were assumed to be similarly low both
forests.
- Paz H. (2003) Root/Shoot Allocation and Root Architecture in
Seedlings: Variation among Forest Sites, Microhabitats, and Ecological
Groups. Biotropica 35(3): 318-332
(abstract) I analyzed patterns variation in root mass allocation and root morphology among
seedlings of woody species in
relation to environmental factors in four Neotropical forests.
Among forests, I explored the response of root traits to
sites varying in water or nutrient availability. Within each forerest,
I explored tha plastic response of species to defferent
microhabitats: gaps and understory. Additionally, I explored evidence for life history
correlation of root and shoot trailts by comparing species differring in their
successional group (light-demanding [22 spp.] or shade torelant [27 spp.])
and germination type (species with photosynthetic cotyledons or species
with reserve cotyledons). At each forest site,
young seedlings from 10 to 20 species were exavated.
A total of 55 species was collected in understory conditions
and 31 of them were also collected in gaps.
From each seedloing, six morphological ratios were determined.
...
Overall, these results suggest that among Neotropical species,
root allocation and root morphology of seedlings reflect plant adjustments to water
or nutrient availability at geographic and microhabitat scales.
In addition, life history specialization to light environments
is suggested by differences among groups of species in their allcation to roots
and in their root morphology.
- Yamada T. et. al (2005) Tap-root depth of trpical seedlings in
relation to species-specific edaphic preferences. Journal of Tropical
Ecology 21: 155-160
(abstract)
...
This trait was fixed independently of the edaphic conditions win whiich the trees
actually grew. Finer-textured soils generally have better water-holding capacity;
we concluded that seasonal water limination would be more severe
in sandy soils than in clay-rich soils, and that water limination would proveke
convergence in seedling root depth
amongh species with the same edaphic preferences.
まあ,
基本的に,
静的な記述で成長過程は調べないんだよね
……
(その場における甲山さんのご指摘なんだけど)
落葉・細根の消失といった「呼吸」のデータはなかなかないからね.
-
まあ,
このあたりの解析すすめるためには,
またいろいろと勉強せねば
……
-
北大構内昼飯調達の旅.
雨はいつのまにかあがっている.
研究室にもどって昼飯.
-
ひさびさに生態学会さーばーネット雑用.
ちょっとアカウント & ディレクトリ作るだけ,
だったんだけど.
-
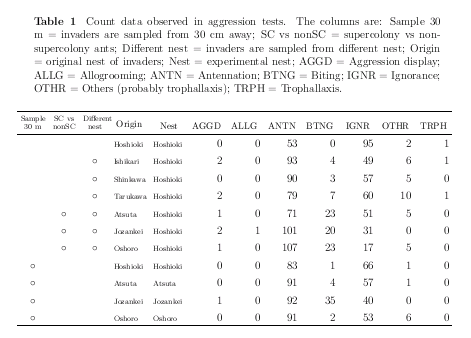
拉致られアリ実験の階層ベイズ推定結果の Table
作り
……
じつは昨日からひっそりと苦闘してる.
うーん,
こんなもんかしらん?
いつものごとく R
& LaTeX コンビネイション自動作表わざ.
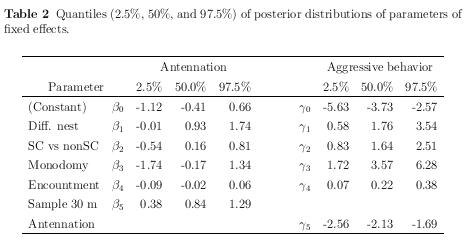
-
カウントデータの Table はレイアウトに苦闘する.
-
2010 研究室発.
2035 帰宅.
体重 67.6kg.
晩飯.
-
明日はなぜかまた甲山さんと
定山渓天狗岳
にのぼりにいくことに
……
-
[今日の運動]
-
[今日の食卓]
- 朝 (0720):
米麦 0.6 合.
コマツナ・マイタケ・卵の炒めもの.
- 昼 (1310):
研究室お茶部屋.
バターロール.
リンゴ.
- 晩 (2200):
米麦 0.8 合.
ネギ・ショウガ・豆腐の炒めもの.
コマツナ・マイタケ・卵の炒めもの.