
[いつのまにか雪]
かとー記念おふぃすからの光景
(東側).
さてセミナー終了後は北大構内走によい時間なんだが
……
けっこう雪がふっている状況.
うんどう不足の日々をすごすうちに冬になってしまったのか.
計算はうまくいってないけど,
だからといって「自分を罰するために雪の中を走る」
という心境にはなれないしなあ.
Moliniopsis-f
(ヌマガヤ開花個体)
で停止しているな
(Trap 窓が出現)
……
集計してみるとたしかにここは 5
個体しかいなくて推定できない,
と.
> sapply(c("s", "j", "f"), function(st)
+ sapply(v.target.spc, function(sp)
+ sum(data[data$spc.stage == paste(sp, st, sep = "-"),]$Freq)))
s j f
Rhynchospora 232 367 287
Hypochaeris 299 180 39
Drosera 153 47 172
Solidago 172 165 13
Moliniopsis 169 132 5
Lobelia 12 153 8
Eriophorum 62 60 1
Carex 17 92 3
s, j, f の 3 段階)
をばらばらに解析するのではなく,
これらをまとめて,
かつ同時に stage 間の相違も推定するよう BUGS 計算まわりを改善する.
6 次元の多変量正規分布を事前分布としてつかわねばならぬ,
と.
cbind(結実した, しなかった)
表記法は使えるは stepAIC()
は使えるは (今日は使わなかったけど),
ホントに便利になりましたねえ.
glmmML()
してみる
……
という部分はささっとできたんだけど,
見ばえのよいテイブルを作るところで苦闘というか,
かなり「汚い」 coding になってしまった.
set AIC autogamy geitonogamy control outcross SD
(naive prob.) (690) 0.08 0.17 0.33 0.51 --
1 (a)(c)(g)(o) 207.9 0.06 0.15 0.31 0.57 0.87
2 (a+g)(c)(o) 212.5 0.10 0.10 0.31 0.57 0.93
3 (a)(c+g)(o) 214.0 0.06 0.22 0.22 0.57 0.95
4 (a)(c+o)(g) 215.8 0.06 0.14 0.44 0.44 0.96
5 (a+g)(c+o) 219.6 0.10 0.10 0.44 0.44 1.01
6 (a+c+g)(o) 229.1 0.15 0.15 0.15 0.58 1.12
7 (a+c)(g)(o) 231.1 0.15 0.14 0.15 0.58 1.12
8 (a)(c+g+o) 233.9 0.06 0.32 0.32 0.32 1.17
9 (a)(c)(g+o) 235.9 0.06 0.33 0.31 0.33 1.16
10 (a+c)(g+o) 251.0 0.15 0.33 0.15 0.33 1.36
...
(a)(c)(g)(o)
が最小になったか.
(naive prob.)
ってのは (結実合計)/(花数合計) という単純なわりざんち.
この AIC が劣悪である理由は
「花序間の overdispersion がとても大きい」
ということだ.
繁殖生態学のヒトたちって,
なぜかしら割算だい好き &
このあたりきちんと解析する気がないんだよねえ
……
glmmML()
とかでやったほうが良いと思うんだが.
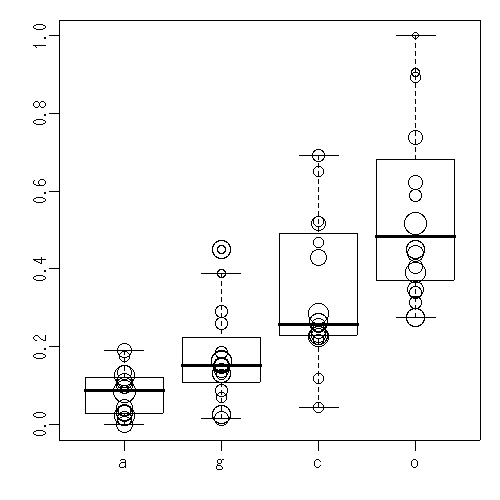
factor(data$treatment, levels = c("a", "g", "c", "o"))
「レヴェル変換わざ」
の実演つき説明.