ぎょーむ日誌 2007-12-02
2007 年 12 月 02 日 (日)
-
0930 起床.
生活周期がよろしくないまま.
あいかわらずうんどう不足な日々だし
……
朝飯.
コーヒー.
怠業.
1120 自宅発.
雪.
1135 研究室着.
-
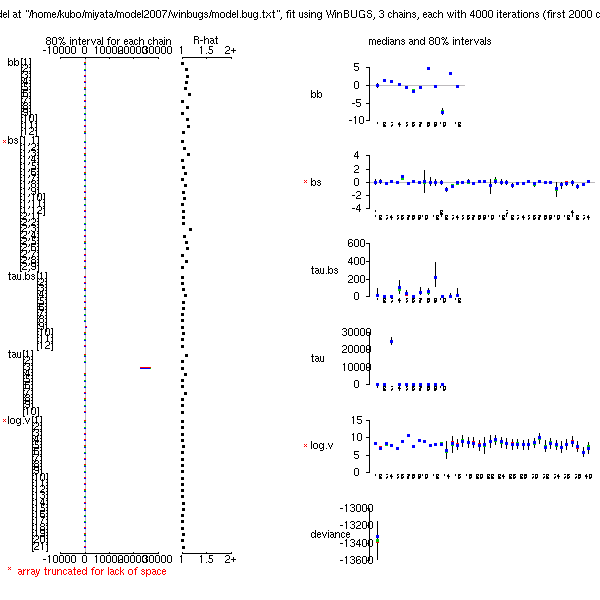
とりあえず昨日の苦闘によりちょー高速化された MCMC 計算の結果
(事後分布表,
BUGS code
……
2007-12-08 追記: これはおおまちがい).
-
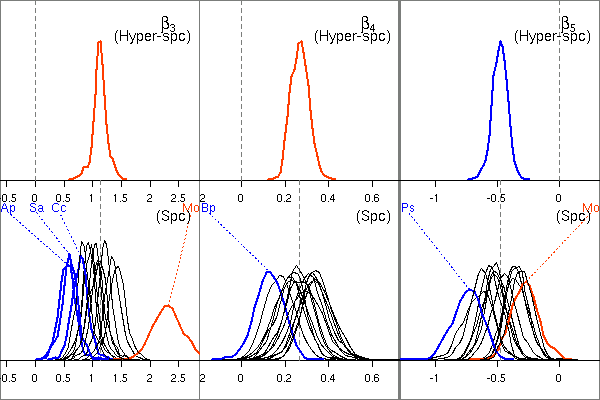
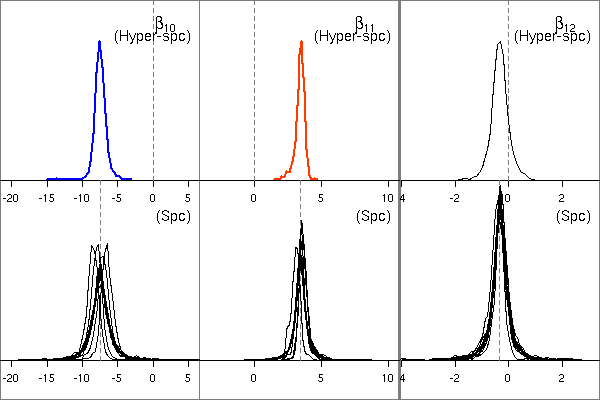
そして体積「分割」パラメーターたちの事後分布図.
個体差 random effects のいれかたがまずいせいか,
「種差」なるものはなさそう
……
あるいはこの観測データからは「種差」なるものが
(もし存在していたとしても)
抽出できないのかもしれない.
-
計算結果の納品作業.
ふーむ,
上記 13 樹種では
長枝・短枝とかいう区別はそれほど明瞭ではなさそう
……
しかし,
樹種によっては何か区別があるようにも見えるし,
その境界線だの短枝出現確率だのが樹高に依存しているのかもしれない.
現時点の統計モデルにはそれが含まれていないけどね.
-
さてさて,
下うけ計算も一段落したことだし査読ぎょーむを
……
と取り組んでみたんだけど,
また熱帯樹冠の階層ベイズモデルの説明責任履行義務が
……
うーむ,
樹木集団の太さ vs 高さ関係なるアヤしげな関係
(これはすごくアヤしい,
と言って悪ければ極端に現象論的なモノのとらえかたです)
を
H = b1 × (1 - exp(-B2 × DBH))
とかいう 2 パラメーターのいいかげんな数式で表現してるんだけど,
「初期における傾き」とやらは
B2
ではなくて
b1 × B2
だなあ
(なんとならば DBH で微分せよ),
ということをど忘れしてた.
いやはや.
-
そして,
その「初期」なる領域ではデータ数がたいへん多いにもかかわらず,
(またしても) 「種差」なるものは見えがたい,
と.
これは「初期」段階で樹木のカタチが良く似ているという意味ではなく,
まったく逆で,
サイズが小さい連中は植物の特性たる可塑性をぞんぶんに発揮してやがるので
カタチは多種多様.
それゆえに,
「種差」なるものが人間には特定しがたい,
という状況なのだろう.
つまり「種差」とかいうものはどうせあるんだろうけど,
あまりにも「個体差」 (これもよくわからぬコトバだが)
が大きいので「種差」なるものはそこに埋没しております,
と.
人間には推定不可能だし
……
そうだな,
群集動態をこねくりまわしている「神の手」からもこぼれおちるだろうね.
-
データを取りまくればますます分類しやすくなるはずだ,
という大 Pearson 的なる信仰は必ずしも正しくないんだよね.
いろいろな状況において.
いっぽうで,
今回の統計モデリングによって観測データが相対的に少ない大サイズ領域では
「種差」らしきものが顕現してるかのように見える
……
まあ,
階層ベイズモデルをもちいることで,
従来方式よりはこのあたりの「データ解析という名のでっちあげ」
が多少はマシになってることでしょうよ.
-
ということで,
よーやく査読ぎょーむに.
うーむ
……
-
うーむ,
で本日はオワってしまって,
2140 研究室発.
雪.
2155 帰宅.
晩飯の準備.
晩飯.
-
[今日の運動]
-
[今日の食卓]
- 朝 (1020):
米麦 0.5 合.
ナス・ピーマン・タマネギ・エリンギ・鶏肉の味噌炒め.
- 昼 (1400):
研究室お茶部屋.
米麦 0.6 合.
春雨スープ.
- 晩 (2250):
米麦 0.8 合.
キャベツ・キュウリ・タマネギのサラダ.
ピーマン・エリンギ・卵の炒めもの.