ぎょーむ日誌 2004-03-05
2004 年 03 月 05 日 (金)
-
0840 起床.
朝からばててる.
0900 自宅発.
晴.
0910 研究室着.
コーヒー.
-
1000 から
論文セミナー,
本日は M2 森君がバルト海アマモの
ヘテロ接合度
(個体内; 遺伝子座数 9)
とクローンサイズの関係を調べた論文を紹介.
調査地によっては heterozygocity とクローンサイズのあいだに
正の相関あるよーなないよーな,
というかんぢなんだが
……
あとから考えてみると,
これって論法としてはけっこうオカルト的なんではなかろうか.
とはいえ,
私のほうも,
いつもながら microsatellite
遺伝子と連鎖の関係をうまく理解できてないんだが.
1120 終了.
-
早起きしたのでばてた.
あるいは計算ばてかもしれん.
メイルやりとり.
お茶部屋に逃げると,
某院生の出身中学校で刀をふりまわす中学生というニュース,
ネットワーク上ではどういう速度で報道されてくか,
という調査につきあってさらにばて.
追出コンパ委員会から何か一筆かけと言われて,
ますますばて.
昼飯.
-
また林冠射影問題にとりくむ.
quadrat
は
corridor
と改名し,
独立したクラス
Corridor
とする.
-
手抜きな近似計算 (Beer 則の誤用もしくは濫用)
する関数かけた
……
ところで,
ナナメからくる光線の光量子密度
(PPFD; µmol m-2 s-1)
を設定しようとするところで,
またつまった.
この「面積あたり (m-2)」って,
ふつーは何の面積をさすのだろうか?
「水平」においた板なのか,
それとも光線と直交する板を考えてるのか
……
谷さんとちょい相談して,
「水平においた板 1m2」
というようなものを念頭におけばよい,
という結論になった.
理由は,
ふつーに使われてる光量子センサーがそういうカタチをしてるから
(で,
センサー内で
µmol m-2 s-1
への換算計算やって出力されてる).
-
しかしこの問題は考えてみると,
そう単純ではない.
仰角 45°に点光源 L があったとしよう.
たとえば
「葉面が L から出てる光線と完全に直交してる場合」
にうけとる光量と 1.0 とする.
この設定で,
もし葉面が光源のほうをむいてるんではなく「真上」
をむいてたとすると
(つまり完全に水平な葉面),
受け取ることのできる光量は 0.71 (= 2-0.5)
ぐらいになってしまう
……
つまりモデルの中ではいろいろな場合について考えんといかん,
ということかね.
-
たとえば,
いま葉群なるものを「球形」としてるけど,
これをたとえば円盤型であっていろんな方向をむいている,
とか
……
まあ,
今回は幸か不幸か間に合いそうにないから,
やらないけどね.
-
じゃあ今回はどうすればよいか?
葉群を球形なんぞと仮定し,
かつ上述のごときいんちきな近似計算をやってる,
というここまでの虚構と矛盾しないウソをでっちあげねばならぬ.
それはすなわち,
葉っぱがホントに球面状に形成されていて,
どんな方向からトんできた光線であっても効率良く受け取れる,
と仮定しつつ受光量を計算すればよい.
すばらしい.
なんで自然界には
これほどまでに優れたボール型葉っぱというものが存在しないのか?
マリモとかそう?
-
ま,
いま考えてるのは葉っぱではなく葉群で,
そのカタマリの中で個々の葉っぱの方向が
……
うんぬん,
という言いわけのほうが無難かな.
いやー,
これはこれでマズいんだよね.
葉群が受け取った光量子が個々の葉っぱに
再配分してやらんといかんから.
とか言いだすと,
一枚の葉っぱであっても光のあたってるところと,
あたってないところで
……
とかいうワケわからん検討に迷いこんでしまう.
植物生態学を研究してるみなさん,
光合成ってのは決定論的な何ものか,
としてとらえたがる傾向があるけれど,
じつはむしろ ``ひどく確率論的'' な現象,
としてパラメーター推定とかせんといかんのぢゃないの?
その生理的詳細が解明されるにつれ.
-
ともかく,
以上の諸点を考慮すれば,
いまの計算において天球光分布を決定するアルゴリズムはこうなるだろう.
-
天球を三角分割していって光源位置をきめる
-
各光源の相対強度
Ri
を決定する;
これは光源の位置と
「晴れてる日」「曇の日」
とかの設定で変わる
(とはいえ,
いまのところ観測データがあまり無さそうなんで,
てきとーに)
-
normalizer を
C = Lstandard
/ (Σ Ri
sinφi)
で計算する;
ただし
Lstandard
は光量子センサーとかで測定される
「その場所の明るさ」
で
φi
は光源 i の仰角
-
各光源の強度を
Li = C Ri
と設定する
こういう計算をやることで,
この数値世界の中であっても
「1 秒間に水平に置いた 1m2 の板に
全方向から飛んできて衝突する光量子の個数」
は 1500 µmol とかになってるわけ
(Lstandard = 1500 とすれば).
-
ふーむ,
やはり
Corridor
内の近似計算方法は
欺瞞能力がびみょーにマズいような気がするな.
まあ,
自分の構築した虚構体系ではこうなってます,
というのが明示的に説明できるようになっているんだから,
それでヨシとするか.
-
しかし全天からの光線うちこみをやる前に,
ある方向からだけの放射について作図してみることにする.
つまり
Corridor なるものの可視化だ.
Perl に特有な ``ad hoc めそっど''
とでも言うような関数を定義していって,
Pov-Ray
にわたす
……
よーし全 Corridor,
光柱を一斉発射,
だ.
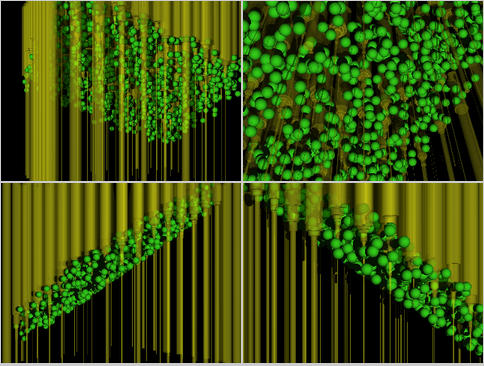
と,
まぁこういうふうになる.
緑球が林冠なるものを構成する葉群である.
黄色い円柱が Corridor
ごとの光量変化を表現している.
葉群にぶつかると光柱が「削られてる」様子がわかる.
Pov-Ray で透過 (transparency) 指定するには
color rgbf <r, g, b, f>.
-
上の図では林冠のほうがナナメになっているのはなぜか
……
それは実際に計算したいのは
「もし斜め上に光源があったら」
という状況で林冠が受けとる光量なんだけど,
これをばか正直にやるよりも,
むしろ逆に
光に関しては常に「垂直うちおろし」,
林冠のほうをぐりぐり回してやったほうが計算がラク,
ということでこういう問題解決になった.
-
さて,
どうして「うちおろし」 (look-down) 式なのか?
以前作ったよーな
「うちあげ」
(look-up)
を使わない理由は,
今回は
「ある方向から流れてこんでくるエネルギー総量」
つまり
Corridor
を走りぬける光子の個数をこちらの思いどーりに統制してやる必要がある.
そのためには「うちおろし」のほうがむいてるからだ.
-
このあたりに関連する計算は来週木曜日の独房群再訪まで,
ある程度やっとかんといかんわけで
……
嗚呼.
-
2100 研究室発.
2120 帰宅.
体重 74.2kg.
晩飯.
-
作図まわりをもう少しヒネる.
そして明日・明後日は夕張で
進化植物学研究会
に参加するんで,
林冠回転計算は今週はここまで.
いやー,
ひさびさの三次元問題,
それも大急ぎでやったんで,
ばてた.
-
[今日の運動]
-
腹筋運動 30 ×
3 回.
腕立ふせ 3 ×
3 回.
-
[今日の食卓]
- 朝 :
食ってない.
- 昼 (1350):
弁当.
研究室お茶部屋.
米麦 0.7 合.
タマネギ・ショウガ・エノキダケ・豆腐の炒めもの.
- 晩 (2230):
米麦 0.7 合.
タマネギ・ショウガ・エノキダケ・豆腐の炒めもの.