ぎょーむ日誌 2001-12-11
2001 年 12 月 11 日 (火)
- 0720 起床.
やや不規則な生活.
- 朝飯・弁当の準備.
朝飯.
シャワー.
- 0750 自宅発.
晴れ.
特急に乗って
0801 京急平和島発.
普通に乗り換えて
0830 京急上大岡発.
0845 研究所着.
- 齡コホートのみ「ニセ」アカマツにおいて,
ぱいぷ逸脱な枝先端部が次第にぱいぴーになる過程を
決定論的モデルとして書き直す作業.
サイズの小さい順に収束速度が一定となるように削減していく
……
というふうにすればいいのかな.
とりあえず割り算を一個減らせる.
- このあたりの元アイデアを以前にご教示してくださった
牧先生
からさらにいろいろとコメントをいただく.
ご提案に対して,
データが無いんで
そんなにパラメーター増やしても推定できません,
と当方の窮状もうしあげると,
ほほほモデリングとはこうするものよと
先生の華麗にして富豪的パラメーター推定法の一端を教えていただいた
……
データがたくさんあったら何をやってもいいんだ,
というカタルシスを体験してしまった.
- 1130 死亡モデルの変更終了.
なんとなくよさそうなカタチには収束する.
しかし枝末端がどんどんアンバランスになっていくような気がするなぁ.
大量生産→大量粛正でむりやりぱいぷな樹木にされるわけだが.
ちょっと極端すぎる.
できごとの順番を少し変えたほうがいいんだけど
……
さて,
ここで観測と矛盾生じないように,
よく考えないといけないんだよね.
- どうしたら良いのかよくわからないので,
とりあえず昼飯.
- 昼からも試運転の続き.
ちょっと細工して「ニセ」アカマツ構成部品が
現在の日本の総人口を超えてもさっさと計算できるようにしてみる.
速い.
しかしなんか変だな
……
- わかった.
Perl であつかえる整数には上限があるんだ.
1569268996
……
15 億 7 千万か.
だいたい毎年 4 倍ぐらいに増えるから
20 世代にいく前にこの数に到達する.
どうしたもんだろ.
- 最初のころはともかく,
いつまでもこの一定速度で増え続ける,
ってのが問題なんだよね.
うう.
それなのに重量は 20 世代で 10-20 倍ぐらいにしか増え
……
うん??
これはどう考えてもばぐだ.
- いつものごとくつまらんミスだった.
毎年毎年あたらしく生成される
RedpineStemCohort
に->SetDensity()
で材密度教えてやるの忘れてた.
まぬけだ.
そして修正プログラムで計算させてみると
……
10 年間で 10 トンの純生産,か.
地球ふろんちあ的に解釈するならば,
こんな妖怪じみた植物が増殖したら
一瞬にして大気中から CO2 が消失して
地球寒冷化するわけだな.
ともあれ,
これで首尾一貫した結果出力になった.
さてどうしたものか.
時刻は 1405.
- 時間とともに各コホートは
ぱいぷ樹木になるという性質をいれてるから
……
根本のほうから猛烈に枯れ上がっているんだけどなぁ.
しかし先端部では指数関数的増殖を食い止められないんだよね.
うーむ.
- 根本方向で枯れが生じたら,
それにあわせて「その先」も落ちるようにしてみるか?
いまでも根本→先端への影響波及はあるんだけど,
徐々にしか伝わらないんだよね.
どういう計算すればいいんだろう.
個々の部品間の親子関係ってのはやめたしなぁ.
- まぁ,
単純に考えて比例配分にするか.
たとえば
下・上ふたつのコホートがあって,
下の面積が 10 で上の面積が 20 とする
(上述のようにゆっくりとしかぱいぴーにならないので,
こういう食い違いは生じうる).
下で 1 死んだらその上では 2 必ず殺す,
と
(さらにぱいぷ教の教えるところに従い
残りの 9:18 という比率が 1:1 に近づいていく).
- 1450 根本→先端への強い波及効果も含めた
死亡モデルができた
……
おお,
ぱいぷ教の呪いが体中をすばやく回ってしまうので
発散するばかりであった樹木部品個数の増殖に
ずずーっとブレイキがかかった.
パラメータいいかげんなんであいかわらず
地球に氷河期をもたらす植物ではあるんだが.
- 収束の時定数を少しあげると強烈にやせ細る
(とはいえまだまだかさばるのだが).
これはなかなか
……
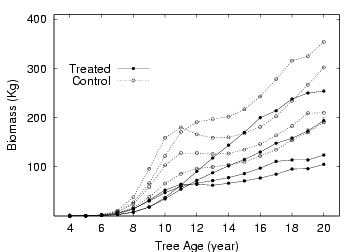
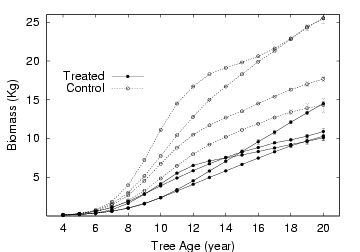
時定数 0.2 とすると個体
2213 の場合,
20 年目あたりでは材乾燥重量が 190Kg ぐらい
(この値はホンモノよりかなり重い?),
部品 (まぁ茎ということで) の個数約 17 万,
なんと 16 万加入して 14.6 万死ぬというような回転なんだよね.
ということで正味 1.4 万個ほどしか増えていない
(重量的にも約 10% の増加).
しかしこの回転率は異常に速すぎるんじゃないのかな.
- 作図でもしてみるか.
このあたりは以前の作図スクリプト (Perl+Gnuplot 連携ワザ)
が使いまわせる.
こういう結果でだいたいいいんだけど
(重量過大だけど)
……
実験区の個体のくせにあとから追いついてくるのがいるな.
こりゃまたどうしてだろう.
- いろいろ調べてみて,
バグでもなさそう,
という結論.
これでもいいんだけどちょっと見てくれが悪い
……
そうだ,
成長曲線を少し曲げてみれば
……
だいたいこんなもんか.
うまくいった.
- なぜ数の変化しか見てない個体群生態学的なモデリングが,
前の (一応はかーぼんばじぇっともどきを計算してみせる)
生産生態学的なモデリングと同じか
それよりマシそうな計算結果を出せるのか.
先のモデリングは観測された個体の枝先を生産器官とみなして,
その個体差だけに着目するものであった.
そういった先端部にも実験処理の影響らしきものは出てるんだけど,
個体によってずいぶんと反応が異なっているので,
その差異をうまくいかしたモデリングは難しい.
- 今回のはいわば個体全体のカタチがどう歪んでるか,
に着目していると言える.
以前に図で示したように,
処理区によってこのパターンが異なっているのである
(実験処理区の樹木たちは根本付近での成長が悪い).
そこでぱいぷもでるという呪法によって,
この歪みを常に個体全体の部品増殖の時間変化に波及させることによって
(成長の悪い部分が一種の「ボトルネック」になっている),
実験区の個体は成長が悪くなるという計算結果を出している.
- 個体全体のカタチのデータを見たときに
「なーんだ,ぱいぷもでるって全然成立してないじゃん」
と考えてたのを
「いや,もし無理矢理にでも成立している,
と仮定するなら樹木はどう成長するだろうか?」
と発想を切りかえたところが反攻の起点になっていたのである
- ……
と対外的にはこういう一見もっともらしい一連の説明でよいのかな.
反攻といっても,全体としては「わー」と敗走中に
ちょっと足を止めて一発喰わせてやった,
ぐらいのもんだし.
これ以上の成果拡大は全く期待できないので,
このへんで落しどころをさぐらなくては.
- 今日はここまで,
か.
バグが残ってるとイヤなんで,
まだ挙動点検せんといかん.
1810 研究所発.
普通に乗って
1820 京急杉田発.
快特に乗り換えて
1830 京急上大岡発.
普通に乗り換えて
1850 京急川崎発.
1920 帰宅.
- また変な時間帯に寝てしまった.
- 今日の食卓
- 朝 (0730):
米 0.6 合.
ナバナ・マイタケの炒めもの.
- 昼 (1205):
弁当.
米 0.8 合.
朝と同じ.
- 晩 (2000):
米 0.6 合.
キャベツ・ニンジン・切り干しダイコンの味噌汁.